
同一把多巴胺鑰匙,為什麼有時讓你「想要更多」,有時卻讓你「徹底厭倦」?
從受體的非線性運算、容積式傳遞到恆定性可塑性,用三個進階層次拆解傳導物質如何在時間中改寫自己。
同一把多巴胺鑰匙,為什麼有時讓你「想要更多」,有時卻讓你「徹底厭倦」?
你已經知道多巴胺(dopamine)編碼的是「獎賞預測誤差」(reward prediction error),也知道受體像鎖、傳導物質像鑰匙。但這裡有一個入門框架解釋不了的現象:一個剛接觸社群媒體的人,每滑出一則新貼文都像中了一次小獎,多巴胺系統興奮地推著他「再滑一下」;可是同一個人連續滑了三小時後,卻陷入一種空洞的、機械式的麻木——明明還在滑,卻一點都不快樂,甚至有點厭惡自己。
同一個分子、同一條迴路,為什麼在幾小時內就從「強力驅動」變成「徒勞空轉」?如果你只停在「多巴胺=動機」這一層,這個轉折是無法理解的。要解開它,我們得把鏡頭從「哪種鑰匙開哪把鎖」,推進到更深的兩個層次:受體被啟動之後,細胞內到底發生了什麼樣的「劑量—反應」運算?以及,當訊號反覆出現時,整個系統如何主動地、有方向地改寫自己?這篇進階篇,就是要帶你進入這兩個層次。
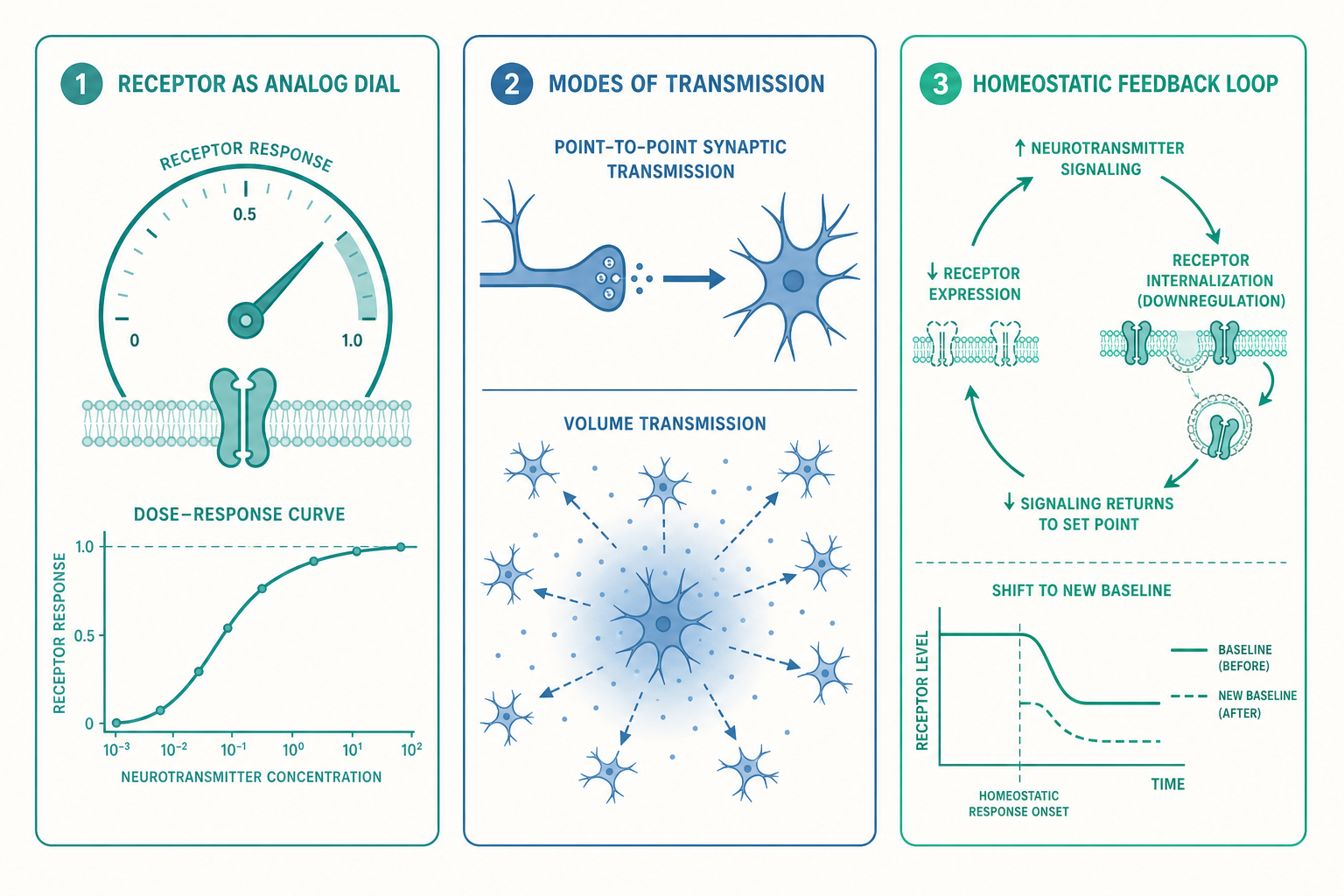
受體不是開關,而是一台類比運算機
入門時我們把受體想成「鎖」——鑰匙插進去,門就開了。這個比喻幫你跨過第一道門檻,但它隱含一個錯誤暗示:受體只有「開」與「關」兩種狀態。事實上,藥理學早在一個世紀前就發現,受體更像一台連續調節的類比運算機,它的輸出由三個彼此獨立的量共同決定。
第一個量是親和力(affinity),描述鑰匙插進鎖的緊密程度,也就是配體(ligand)與受體結合的牢固度。第二個量是效能(efficacy),描述鑰匙轉動後「能把門推開多少」。這兩者必須分開看,因為存在一類關鍵的分子——拮抗劑(antagonist):它親和力很高、牢牢佔住受體,效能卻是零,等於一把插進鎖孔卻轉不動、還擋住真鑰匙的假鑰匙。很多藥物正是靠這種「高親和、零效能」來阻斷訊號。
更微妙的是第三個量。傳統觀念認為一個受體只有一種「啟動」方式,但 1990 年代後確立的偏向性致效(biased agonism / functional selectivity)徹底改寫了這個圖像。原來,一個代謝型受體(metabotropic receptor,多屬 G 蛋白偶聯受體,G protein-coupled receptor, GPCR)被啟動後,下游同時連著好幾條路徑——例如 G 蛋白路徑與 β-arrestin 路徑。不同的配體能讓受體「偏向」啟動其中某一條,而非全部。這意味著:同一個受體,被不同分子啟動時,會送出質地不同的訊號。 鑰匙與鎖的比喻到這裡需要升級——鑰匙不只決定「開哪扇門」,還決定「門開之後,屋裡哪幾盞燈會亮」。這正是新一代止痛藥研發的核心策略:能否找到只啟動類鴉片受體止痛路徑、卻不啟動成癮與呼吸抑制路徑的「偏向性配體」。
看一個例子:用「佔據率」算給你看
把上面的話量化,你會更有感。藥理學常用一個簡化的佔據率(occupancy)關係:當配體濃度等於它的解離常數(dissociation constant, K_d,數值越小代表親和力越高)時,恰好有一半的受體被佔據;濃度提高到 K_d 的十倍時,約佔據九成;再提高十倍,約佔據九成九。
注意這條曲線的形狀:它不是直線,而是先陡後平的飽和曲線。這帶來一個極其重要、卻常被忽略的後果——當受體已經接近飽和,再增加傳導物質濃度,效果幾乎不再增加。 這就解釋了為什麼很多精神科藥物存在「天花板效應」:劑量加倍,療效卻沒有加倍,副作用反而先到。它也回頭補強了入門篇 SSRI 的故事:單純把突觸血清素濃度推高,並不會線性地把心情推好,因為受體端的回應本就是飽和的、非線性的。理解傳導物質「量的多寡」,一定要同時理解受體端這條「非線性回應曲線」,兩者缺一不可。
點對點,還是廣播?兩種截然不同的傳遞模式
入門篇描繪的突觸傳遞,是一幅乾淨俐落的「點對點」圖像:一個神經元對準另一個神經元,把分子精準投遞到二十奈米寬的縫隙裡。這叫接線式傳遞(wiring transmission),麩胺酸(glutamate)與 GABA 的快訊號大致如此——它們追求的是速度與精準,毫秒級、一對一。
但多巴胺、血清素、正腎上腺素(norepinephrine)這些「神經調節物質」(neuromodulator),玩的卻是另一套規則,稱為容積式傳遞(volume transmission)。它們釋放後不急著被回收,而是擴散到突觸結構之外,瀰漫在一片組織液裡,去影響附近成百上千個並沒有與它直接接線的神經元。這不是點對點的私訊,而是對整個街區廣播的擴音器。
這個區分為什麼重要?因為它從根本上決定了一種傳導物質的「功能身份」。接線式傳遞傳遞的是內容——具體是哪個神經元在何時放電;容積式傳遞傳遞的是脈絡(context)——整片迴路此刻處於什麼樣的全域狀態:是警覺還是放鬆?是探索還是利用?是該相信舊習慣還是該嘗試新策略?這也說明了為什麼神經調節物質的作用往往慢、廣、而深遠,且難以用單一突觸的實驗去捕捉。當你讀到「血清素影響情緒」時,真正的機制不是某條線路被接通,而是大片皮質的運作背景被悄悄調了一個檔位。
增益調節:神經調節物質真正在運算什麼
那麼,「調背景」具體是調什麼?當代計算神經科學給出一個強而有力的答案:增益調節(gain modulation)。
想像每個神經元都有一條「輸入—輸出」轉換曲線(一般是 S 形):輸入訊號越強,輸出放電率越高,但到頂後飽和。神經調節物質的核心作用之一,就是改變這條曲線的陡峭程度,也就是增益。增益調高,神經元對輸入差異變得敏感、反應銳利、訊噪對比拉大;增益調低,反應變得平緩、遲鈍、趨於隨機。換句話說,它們不直接決定「答案是什麼」,而是決定「整個系統此刻有多認真看待輸入」。
正腎上腺素是這套理論最清楚的範例。Aston-Jones 與 Cohen 在 2005 年提出影響深遠的理論,主張腦幹藍斑核(locus coeruleus)釋放的正腎上腺素,調節的正是大腦在「利用(exploit)既有策略」與「探索(explore)新可能」之間的切換。低而穩定的正腎上腺素背景,讓系統專注、堅持當前任務;陣發性的高正腎上腺素,則打散當前狀態、開放系統去嘗試別的選項。你會發現,這跟強化學習(reinforcement learning)裡著名的「探索—利用權衡」(exploration-exploitation tradeoff)是同一個問題的兩種語言——一邊是腦幹的化學廣播,一邊是演算法的超參數。這條跨領域的對應,正是 AI 與神經科學持續對話的沃土。
血清素則被認為扮演著與多巴胺互補、甚至「對立」的角色。一條有力的假說(由 Daw、Dayan 等人發展)主張:若多巴胺主管對「獎賞」的學習,血清素可能主管對「懲罰、損失與等待」的處理,調節我們願意為了遠期回報而抑制衝動、忍耐當下的能力。這也比「血清素=幸福分子」精確得多——它管的不是幸福本身,而是我們如何在時間維度上權衡苦與樂。
系統會反咬自己:耐受、戒斷與成癮的分子邏輯
現在我們回到開篇那個謎題:為什麼滑手機三小時後,多巴胺驅動的快感會塌縮成空洞的麻木?答案藏在一個入門篇刻意留白、卻是整個神經科學最深刻的原理之一——恆定性可塑性(homeostatic plasticity)。
神經系統有一個近乎執拗的傾向:它討厭任何訊號長期維持在異常高或異常低的水平,並會主動反向調整,把活動拉回它認定的「設定點」。當某條迴路被反覆、過量地刺激(無論是成癮藥物、還是設計成讓你欲罷不能的數位產品),突觸後神經元會做出一系列代償:把細胞膜上的受體「內吞」收進細胞裡,減少受體數量(受體下調,receptor downregulation);或降低剩餘受體的敏感度。結果是,同樣強度的刺激,能引發的反應一次比一次小。 這就是耐受性(tolerance)的分子基礎,也是為什麼第一根菸、第一次滑手機的衝擊,再也回不來。
更殘酷的是這套代償的「反作用」。當系統已經把受體調降、把基準線往下壓,一旦外來刺激突然撤除,剩下的內生訊號便嚴重不足——這就是戒斷(withdrawal)的負面情緒與身體不適。成癮的核心痛苦,很大一部分並非來自「得不到快感」,而是來自這套被改寫過的恆定系統留下的、低於正常的新基準線。神經科學家 Nesler 等人發現,長期藥物暴露還會累積一種異常穩定的轉錄因子 ΔFosB,它像一個分子層級的「記憶痕跡」,長期改寫基因表現,被認為是成癮容易復發、難以根治的部分原因。
把這條線索接回優心理學,你會看到行為層次的「習慣化」「報酬遞減」「衝動控制」,在分子層次都有著對應的硬體基礎。也把它接回 AI:當推薦演算法不斷餵給你「比預期更好一點點」的內容,它其實是在系統性地利用你大腦的獎賞預測誤差機制——而你的恆定性可塑性,則註定了這場互動會走向耐受與空虛。理解這套機制,不是為了恐嚇,而是為了讓你在面對被精心設計來「劫持」這套化學語言的產品時,多一分清醒的自主。
動手試試:用三層框架拆解一個成癮迴圈
不需要任何設備,試著用今天的三個進階工具,分層拆解「短影音上癮」這個現象:
-
受體運算層。 每一則新影片帶來一次多巴胺脈衝。問自己:隨著一次次刺激,突觸後受體的「佔據率—回應曲線」會如何被恆定性可塑性往哪個方向推?(提示:受體下調,曲線整體右移、變鈍。)
-
傳遞模式層。 多巴胺在這裡走的是容積式還是接線式傳遞?這對「你很難只想看一支就停」這件事有什麼解釋力?(提示:被調的是整片迴路的全域狀態,不是某條精準線路,所以難以靠意志精準關掉。)
-
增益與探索層。 用 Aston-Jones–Cohen 的框架想想:無止盡的新鮮刺激,可能讓系統長期卡在哪一種模式(探索還是利用)?這跟你「滑了三小時卻什麼都沒記住」有沒有關係?
做完這個練習,你會發現自己已經能在同一個現象上自如地切換三個分析層次。能做到這件事,正是進階學習者與入門者的分水嶺。
重點回顧
- 受體是類比運算機,不是開關:輸出由親和力、效能、偏向性致效三者共同決定;佔據率曲線是非線性飽和的,故有「天花板效應」。
- 兩種傳遞模式承載不同資訊:接線式傳遞(麩胺酸、GABA)傳「內容」、快而精準;容積式傳遞(多巴胺、血清素、正腎上腺素)傳「脈絡」、慢而廣泛。
- 神經調節的核心運算是增益調節:它不決定答案,而決定系統此刻多認真看待輸入;正腎上腺素對應探索—利用切換,血清素可能對立於多巴胺、主管損失與耐心。
- 恆定性可塑性讓系統反咬自己:反覆刺激引發受體下調,造成耐受;撤除後基準線過低,造成戒斷——這是成癮痛苦的分子核心。
- 量與受體回應、傳遞模式、系統可塑性必須一起看:單看「某物質多了或少了」永遠無法解釋真實的動態行為。
深入探討(研究所視角)
走到這裡,你手上已經有了一套遠比「快樂分子/幸福分子」精細的分析語言。研究所層次的關鍵,是把這些片段整合成一個動態系統控制的整體觀,並學會質疑它的邊界。
相位與張力:同一物質的兩種時間碼。 多巴胺神經元有兩種放電模式——緩慢持續的「張力性(tonic)」基準活動,與快速陣發的「相位性(phasic)」尖峰。前者設定整體動機的「水位」,後者承載我們熟悉的獎賞預測誤差訊號。Grace 等人的研究顯示,這兩種模式由不同機制調控,且在思覺失調症(schizophrenia)等疾病中可能各自失常。這帶出一個對研究者極重要的方法學警示:你在資料中看到的「多巴胺活動」,到底測的是張力性水位、還是相位性尖峰?兩者混為一談,會導出截然相反的結論。同一個分子,在不同時間尺度上編碼著完全不同的變數——這是計算神經科學最迷人也最棘手的特徵之一。
從單一誤差到分佈式預測。 入門篇提到多巴胺實現了時間差分學習(TD learning)的誤差項 δ。但 2020 年 Dabney、Kurth-Nelson 等人在《Nature》發表的研究更進一步:他們發現中腦多巴胺神經元並非全都編碼同一個平均化的預測誤差,而是各自有不同的「樂觀/悲觀」偏好,整體上實現了 AI 中的分佈式強化學習(distributional reinforcement learning)——大腦編碼的不是「期望報酬」這個單一數字,而是報酬的整個機率分佈。這是近年神經科學與機器學習雙向啟發的範本:一個源自 AI 的演算法改良,反過來預測並被證實於真實神經元的活動。它呼應了 Uedu 人機共學的精神——理解大腦如何學習與理解機器如何學習,本就是同一場探索的兩面。
膠細胞與細胞外環境:被遺忘的第三方。 經典模型只談神經元,但越來越多證據顯示,星形膠細胞(astrocytes)不是被動的填充物——它們會回收麩胺酸、釋放「膠傳遞物質」(gliotransmitter)、主動參與突觸的調節(「三方突觸」,tripartite synapse 概念)。把膠細胞納入,傳導物質的故事才算完整。這也是當前最活躍的爭論前沿之一。
跨領域連結與開放問題。 傳導物質研究天然橫跨多個學門:往下接優生物(GPCR 的晶體結構、β-arrestin 訊號的結構基礎、轉錄因子如何重塑突觸);往上接優心理(成癮、衝動、跨期決策的行為模型);橫向接 AI(分佈式 RL、增益調節作為網路超參數、探索—利用權衡)。當前最值得追問的開放問題包括:能否用「增益/神經調節狀態」這種系統層次的標記,取代過時的「單一分子失衡」來理解精神疾病?偏向性致效能否真的兌現「有療效、無成癮」的止痛藥承諾?以及——這也直接連到 Uedu 的 PhysioNeuromics 維度——若神經調節狀態(警覺、探索傾向、增益高低)會外顯於 HRV、瞳孔大小、睡眠等生理多模態訊號,那麼我們能否從學習者的生理資料,間接推估其當下的神經調節狀態,進而為「什麼時候該給挑戰、什麼時候該給鞏固」這類學習設計,提供有原理依據的時機判斷?這些問題都還沒有定論,正等待用比「快樂分子」更精確的語言去回答它們的新一代研究者。