
為什麼「想起來」這個動作,反而可能改寫你的記憶?
深入記憶的時間維度:系統固化的腦區搬家與理論之爭、提取誘發的再固化編輯窗口、睡眠重播的神經耦合,以及為何遺忘是一種適應性設計。
為什麼「想起來」這個動作,反而可能改寫你的記憶?
入門篇結束在一個令人安心的圖像:經驗透過長期增益(long-term potentiation, LTP)被刻進突觸權重,再經由睡眠固化成長期記憶,彷彿一旦寫入便永久封存。但這裡有個違反直覺的事實,是當代記憶研究最迷人的轉折之一:每一次你成功「提取」(retrieve)一段記憶,那段記憶都可能短暫地變回不穩定狀態,需要重新被固化一次。 換句話說,回憶不是唯讀的播放,而更像把檔案打開來編輯——你有機會增補、扭曲,甚至(在特定條件下)抹除它。
這篇進階文章不再重複「記憶如何形成」,而是聚焦三個入門篇刻意略過的時間維度難題:(1) 記憶形成之後,如何在海馬迴與大腦皮質之間「搬家」與「轉型」(系統固化的兩大對立理論);(2) 提取為何會讓記憶再度可塑(再固化,reconsolidation);(3) 睡眠中的神經重播(replay)究竟在分子與系統層級做了什麼。最後我們會看到,這些機制如何重新定義「遺忘」,並與人工智慧的記憶難題交會。
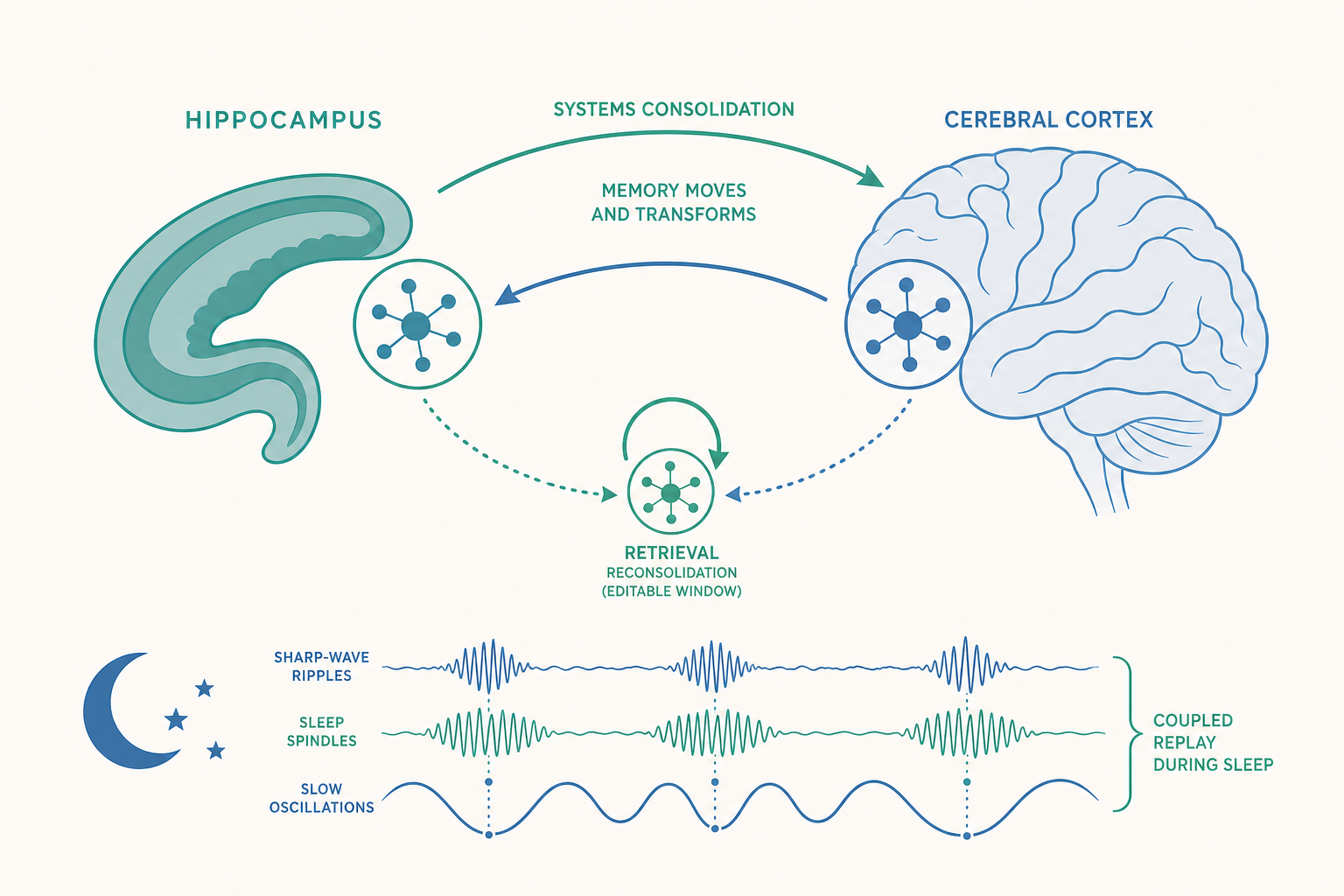
系統固化:記憶為什麼要「離開」海馬迴?
入門篇談的 LTP、NMDA 受體、CREB,都發生在單一突觸或單一神經元尺度,時間跨度是秒到天。但記憶還有一個更大尺度的故事:系統固化(systems consolidation),跨越的是數週、數月甚至數年,談的是記憶在不同腦區之間的重新分配。
證據的起點是一位代號 H.M. 的病人(Henry Molaison)。1953 年,為治療嚴重癲癇,外科醫師切除了他兩側的內側顳葉,包含大部分海馬迴。手術後,H.M. 的智力、語言、工作記憶都正常,舊有的童年記憶也大致完好——但他再也無法形成新的長期外顯記憶。更關鍵的線索是:他遺忘的並非所有舊記憶,而是手術前幾年的記憶受損最重,越久遠的越完整。這種「時間梯度的逆行性失憶」(temporally graded retrograde amnesia)強烈暗示:海馬迴是新記憶的暫時樞紐,但記憶會隨時間逐漸「獨立」於海馬迴之外。
標準固化模型 vs 多重痕跡理論
這個現象催生了兩個彼此競爭的理論,至今仍是研究所考題的常客:
- 標準固化模型(Standard Consolidation Model, SCM):海馬迴像一位臨時的「索引器」。剛學會時,記憶內容分散儲存在皮質各處,但要靠海馬迴把它們綁在一起、一同喚起。隨著時間(特別是睡眠中的反覆重播),皮質神經元之間直接建立連結,記憶逐漸「皮質化」,最終不再需要海馬迴。這解釋了為何久遠記憶在海馬迴受損後仍存在。
- 多重痕跡理論(Multiple Trace Theory, MTT),由 Moscovitch 與 Nadel 提出:他們指出,情節記憶(episodic memory,帶有具體時空脈絡的個人經歷)永遠需要海馬迴。每次提取一段情節記憶,就會在海馬迴留下一個新的痕跡,使舊記憶擁有多重備份,因而對海馬迴局部損傷更有抵抗力。只有抽離了脈絡、變成「語意化」(semantic)的一般知識,才會真正獨立於海馬迴。
兩者的核心分歧在於:老舊的情節記憶到底還需不需要海馬迴? SCM 說不需要,MTT 說情節記憶永遠需要。神經影像研究發現,回憶生動、細節豐富的久遠往事時,海馬迴仍會活化,這對 MTT 較有利;但臨床上某些海馬迴受損病人確實保有久遠記憶,又支持 SCM。目前較被接受的整合觀點是「軌跡轉型理論」(Trace Transformation Theory):記憶不是單純地「搬家」,而是隨時間轉型——細節豐富、依賴海馬迴的情節版本,與抽象、依賴皮質的要旨(gist)版本可以並存,端看你提取的是哪一種。
這個區分對學習者極有意義:你準備考試時想記住的「概念要旨」,與你日後回想「那堂課發生了什麼」的生動情節,走的是不同的固化路徑、依賴不同腦區。
再固化:提取打開了一扇「編輯視窗」
如果固化是「把記憶寫穩」,那再固化(reconsolidation)就是「記憶被讀取後,必須重新寫穩一次」。
這個概念的現代復興來自一個漂亮而簡潔的實驗邏輯(Nader、Schafer 與 LeDoux, 2000)。研究者先讓大鼠形成一段恐懼記憶(聲音搭配電擊)。隔天,他們在大鼠的杏仁核(amygdala,恐懼記憶的關鍵腦區)注入蛋白質合成抑制劑。重點在於注入的時機:
- 如果只注入抑制劑、不喚起那段記憶,記憶完好如初。
- 如果先播放那個聲音、喚起記憶,再注入抑制劑——那段原本已經穩固的記憶,竟然消失了。
這顛覆了「固化只發生一次」的經典觀點。它意味著:提取會把記憶帶回一個不穩定、可塑的狀態,這時若阻斷蛋白質合成,記憶就無法被「重新寫穩」,於是流失。記憶被讀取的那一刻,短暫地變回了「可編輯文件」。
看一個例子:用一個生活情境理解再固化
假設你對某次公開報告留有強烈的焦慮記憶。每次你回想那次經驗(提取),記憶就進入可塑窗口。若這時你伴隨的是新的、緩和的脈絡——例如在放鬆狀態下重新講述、得到正向回饋——那麼當記憶「重新寫穩」時,被固化下來的就是一個略微更新、焦慮成分稍弱的版本。反過來,若你每次回想都更加緊張,再固化反而會強化焦慮。
這正是「暴露療法」(exposure therapy)與一些創傷治療新取向的理論基礎之一:不是抹除記憶,而是在記憶被提取、進入可塑狀態時,趁機更新它的情緒與意義成分。Schiller 等人(2010)甚至證明,在人類身上,於再固化窗口內進行行為層級的消退訓練,能持久削弱恐懼反應——完全不需藥物。
對日常學習而言,再固化也提供了一個重要洞見:每一次檢索練習(retrieval practice)都不是中性的「測試」,而是一次改寫記憶的機會。 你提取得越正確、提取時建立的連結越豐富,重新固化下來的記憶就越強、越有彈性。這從機制層面解釋了為何「考試效應」(testing effect)——主動回憶比反覆閱讀更有效——如此可靠。
睡眠重播:海馬迴在夜裡「快轉」白天
入門篇提過睡眠有助記憶固化,但機制值得深入。1994 年,Wilson 與 McNaughton 在大鼠身上做出了一個標誌性發現:白天當大鼠在迷宮中走過某條路徑時,海馬迴會有一組「位置細胞」(place cells)依特定順序放電;當晚大鼠進入睡眠(特別是慢波睡眠,slow-wave sleep)時,同一組細胞會以相同的順序、但壓縮約 10–20 倍的速度,自發地重新放電一遍。 這就是神經「重播」(replay)。
重播為什麼重要?因為它正是系統固化的生理引擎。在慢波睡眠中,三種腦電節律精準耦合:
- 海馬迴的尖波漣漪(sharp-wave ripples, SWRs):重播事件就嵌在這些短暫的高頻爆發中。
- 丘腦皮質的睡眠紡錘波(spindles)。
- 皮質的慢振盪(slow oscillations)。
當這三者在時間上巧妙嵌套,海馬迴的重播訊號便能在皮質「放下訊號」的最佳時間窗,反覆地對皮質「教學」,逐步把記憶轉移過去。實驗證據相當直接:以電刺激人為打斷大鼠的 SWRs,會選擇性地損害空間記憶的固化,但不影響其他能力。換言之,沒有重播,固化就跛腳。
更精緻的是,重播並非單純複製。睡眠中的重播會偏好白天帶有獎勵或情緒標記的經驗,也會把不同片段重新組合、推演從未實際走過的捷徑路徑。這暗示睡眠不只是「存檔」,還在做整理、抽象與推論——這正呼應了系統固化裡「情節 → 要旨」的轉型。
動手試試:替自己設計一個「機制一致」的讀書計畫
試著只用本文與入門篇的機制,推導出一份讀書計畫,並對每一步寫出對應的神經理由:
- 學完新內容後,安排一段充足睡眠再進入下一章——理由:慢波睡眠的 SWR 重播驅動系統固化,犧牲睡眠等於關掉固化引擎。
- 複習時用「闔上書本主動回憶」取代「重讀」——理由:提取把記憶帶入再固化窗口,正確提取會重新寫穩並強化記憶(考試效應)。
- 把複習分散在數天,每次間隔拉長——理由:間隔讓每次提取都重新觸發固化、推進到需要蛋白質合成的晚期 LTP,並讓痕跡逐步皮質化。
- 回憶時刻意連結其他已知概念、舉自己的例子——理由:再固化會把提取當下建立的新連結一併寫入,使要旨版本更豐富、更獨立於原始脈絡。
如果你能對每一步都說出機制理由,代表你已能把分子、系統、行為三個層級串成一條因果鏈——這正是研究所層級對「學習科學」的要求。
重新理解「遺忘」:不是失敗,而是設計
把上述機制整合起來,遺忘的面貌就完全改變了。它至少有三種彼此不同的形式:
- 固化失敗:記憶根本沒推進到晚期 LTP 與系統固化,停在脆弱的早期階段就消散。臨時抱佛腳大多屬此。
- 提取失敗:記憶痕跡仍在,但缺乏有效的提取線索(cue)把它喚出。這解釋了「舌尖現象」與「換個情境就想起來」。
- 主動抑制與更新:透過 LTD(長期抑制)的突觸修剪、再固化時的覆寫、以及提取誘發的遺忘(retrieval-induced forgetting,提取某項記憶會抑制相關競爭記憶),大腦主動地讓無關訊號退場。
關鍵觀念是:遺忘往往是適應性的(adaptive forgetting)。 一個記得每一個細節、無法遺忘的大腦(如某些「超憶症」hyperthymesia 案例所顯示)並不見得更聰明,反而可能被無關細節淹沒、難以抽取要旨與類化。學習的目標從來不是「記住一切」,而是讓重要的訊號從雜訊中浮現、並能彈性地遷移到新情境。從這個角度看,良好的遺忘與良好的記憶同樣是學習的成就。
重點回顧
- 記憶會在腦區之間「搬家並轉型」:系統固化讓記憶逐步從依賴海馬迴轉向皮質;標準固化模型與多重痕跡理論的爭論,核心在於老舊情節記憶是否仍需海馬迴,整合觀點(軌跡轉型理論)認為細節版與要旨版可並存。
- 提取會讓記憶再度可塑:再固化現象顯示,喚起一段記憶會把它帶回不穩定狀態,需要重新合成蛋白質才能寫穩——這是更新乃至治療性改寫記憶的窗口。
- 檢索練習之所以有效,有分子層級的原因:每次成功提取都是一次再固化,會強化並更新記憶,這是「考試效應」的機制基礎。
- 睡眠重播是固化的生理引擎:慢波睡眠中海馬迴尖波漣漪、睡眠紡錘波與皮質慢振盪三者耦合,把白天經驗壓縮重播並轉移到皮質;打斷漣漪會選擇性損害固化。
- 遺忘是多種機制、且常是適應性的:固化失敗、提取失敗與主動更新各不相同;能遺忘無關訊息、保留要旨,本身就是健康學習的一環。
深入探討(研究所視角)
再固化的「邊界條件」與臨床轉譯的現實。 再固化聽起來像是「改寫創傷記憶」的萬靈丹,但研究很快揭示了嚴格的邊界條件(boundary conditions)。並非每次提取都會開啟可塑窗口:記憶必須夠新、提取時必須出現「預測誤差」(prediction error,即現實與預期不符的訊號),窗口才會打開;過於陳舊或過度強化的記憶,可能進入「不再固化」的抵抗狀態。這背後可能涉及不同分子開關(如 GluN2B 次單元 NMDA 受體、以及一系列調控蛋白)。對學習者的啟示是:別把單一動物實驗的乾淨結論直接外推到複雜的人類臨床或教學現場——機制為真,但條件決定成敗。這也是評讀文獻時最該追問的:「這個效果在什麼邊界內成立?」
從突觸到細胞群體:engram 在固化中如何重組。 結合再固化與系統固化的前沿問題是:當記憶從海馬迴轉移到皮質,engram 細胞集合本身發生了什麼? Kitamura、Tonegawa 等人(2017)用標記技術發現,皮質的 engram 在學習當下其實就已「沉默地」生成(silent engram),需要海馬迴的反覆驅動數週後才「成熟」並能獨立被自然線索喚起。這意味著系統固化不是把記憶從一處複製到另一處,而是讓一個原本沉默的皮質痕跡逐漸「上線」。這與互補學習系統(complementary learning systems)理論高度契合,也讓「記憶到底儲存在哪裡」的答案變成「同一段記憶在不同時間,主要由不同的細胞集合承載」。
跨領域連結——AI 的災難性遺忘與「人造睡眠」。 入門篇提過 Hebbian learning 是人工神經網路的源頭,這裡可以接得更深。人工神經網路面對「災難性遺忘」(catastrophic forgetting):學新任務時權重被覆寫,舊任務隨即崩潰——這正是缺乏系統固化的後果。有趣的是,AI 工程師借鏡大腦的解法竟與睡眠重播高度相似:經驗回放(experience replay) 讓模型反覆「重播」儲存的舊樣本,正對應海馬迴的 SWR 重播;生成式重播(generative replay) 用一個生成網路「做夢」般地合成舊任務的假樣本來複習,對應睡眠中重播的重組與推演特性;而 elastic weight consolidation 等演算法則模仿突觸鞏固,對重要權重「上鎖」以防覆寫。這條從 H.M.、位置細胞重播,一路通到 AI 持續學習(continual learning)的線索,正是 Educational Omics 中 Cognomics(認知歷程)與當代人工智慧最深刻的交會點之一。
回到課堂的整合視角。 把本文四條主線——系統固化的腦區轉移、提取誘發的再固化、睡眠重播的生理耦合、適應性遺忘——疊在入門篇的分子與突觸機制之上,學習者會得到一個遠比「多複習就會記得」更立體的圖像:記憶是一個動態、會隨時間轉型、且在每次回憶時被重寫的歷程。許多被實證反覆驗證的策略——間隔練習、檢索練習、充足睡眠、把新知連結到舊知——之所以有效,是因為它們各自精準地搭上了某一條生物時程。當你理解大腦不是被動地「儲存」經驗,而是主動地建構、轉型與更新記憶,你就握有了與自己大腦更聰明合作的鑰匙——也更能用批判的眼光,去評估任何宣稱能「強化記憶」的工具或療法究竟踩在哪一條機制之上。