
如果軸突可以長到一公尺,細胞核要怎麼「指揮」一公里外的工地?
從軸突運輸、離子通道時序到軸突起始段與樹突分區運算,看神經元的構造如何成為一個動態的訊號處理系統。
如果軸突可以長到一公尺,細胞核要怎麼「指揮」一公里外的工地?
想像你是一家工程公司的總部,所有的設計圖、所有的建材都只能從你這裡發出,但你的工地遠在一公尺之外——以細胞的尺度換算,這相當於一個人要遠端管理一座一千公里外、沒有當地分公司的建築工程。這正是一個運動神經元(motor neuron)每天面對的處境:它的細胞核安坐在細胞本體裡,而它的軸突末梢卻可能遠在腳趾。蛋白質只在細胞本體合成,囊泡也多半在那裡組裝,可是訊號的釋放、膜的修補、能量的供應卻全發生在遙遠的末端。
入門篇我們把神經元拆成樹突、細胞本體、軸突、突觸四個部位,講清楚了「接收—整合—輸出—傳遞」的分工。但那是一張靜態的解剖圖。一旦你問「這些構造怎麼維持、訊號怎麼產生、運算**到底發生在哪裡」,神經元就從一張圖變成一個被精密物流、離子電化學與分區計算共同支撐的動態系統。這一篇,我們就鑽進這些被入門略過的機制裡。
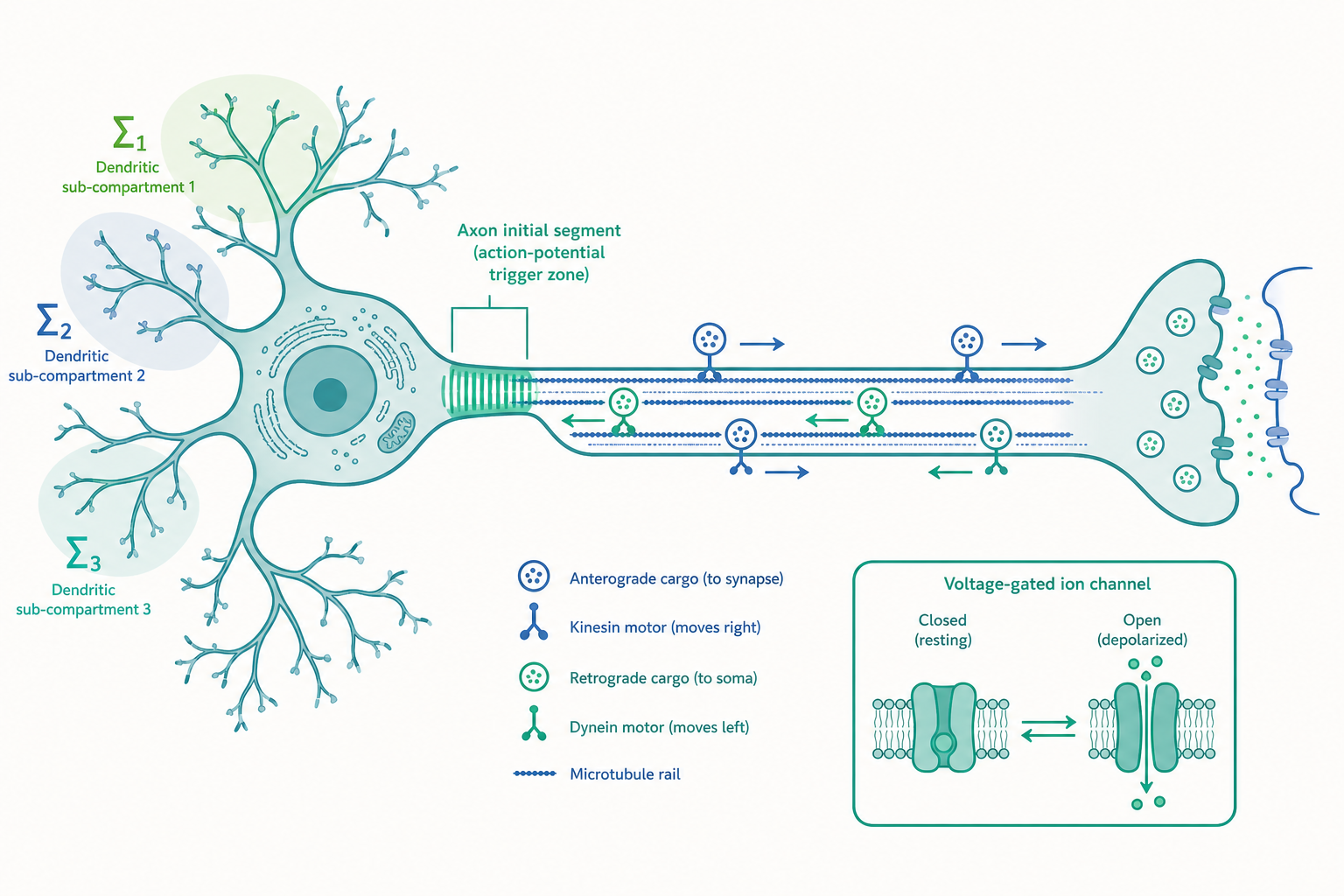
軸突運輸(Axonal transport):細胞內的物流幹線
一個神經元最反直覺的事實是:它的體積有時九成以上落在軸突,但合成蛋白質的機器(核糖體、內質網、高基氏體)幾乎都擠在細胞本體。這代表末梢需要的所有「貨物」——突觸囊泡的前驅物、離子通道、線粒體、訊息分子——都得從總部一路運過去。負責這條物流幹線的,是軸突內部一束平行排列的微管(microtubule),它是細胞骨架的一部分,本身具有方向性:朝向細胞本體的一端稱為負端,朝向軸突末梢的一端稱為正端。
兩種馬達蛋白沿著微管走相反的方向。驅動蛋白(kinesin) 把貨物從細胞本體往末梢運(順向運輸,anterograde),動力蛋白(dynein) 則把訊息與待回收物從末梢運回細胞本體(逆向運輸,retrograde)。它們像在鐵軌上行走的搬運工,靠水解 ATP 一步一步「邁步」前進。順向的快速運輸速度可達每天數百毫米,慢速運輸(搬運細胞骨架蛋白與可溶性酵素)則只有每天數毫米。
這條幹線一旦故障,後果嚴重。許多神經退化疾病的共同特徵就是軸突運輸受損:當馬達蛋白、微管或被運送的貨物(例如線粒體)出問題,遠端的突觸會先「斷糧」而退化——這種「由末梢往內死亡」的模式稱為逆死性退化(dying-back),在肌萎縮性脊髓側索硬化症(ALS)等疾病中可見。逆向運輸還有一個我們每天都依賴的功能:把末梢偵測到的「環境狀態」(例如標的細胞分泌的神經滋養因子 NGF)回報給細胞核,讓細胞核決定基因該怎麼表現。換句話說,總部之所以知道一公尺外工地的近況,靠的正是這條逆向回報的訊息鏈。
動作電位的真正引擎:離子通道的開與關
入門篇說動作電位是「全有全無」、靠頻率編碼強弱。但它為什麼能做到全有全無?答案藏在膜上電壓門控離子通道(voltage-gated ion channel) 的開關時序裡,而這正是 Hodgkin 與 Huxley 在 1952 年用槍烏賊巨大軸突解開的謎題。
靜止時,神經元膜內外維持約 −70 毫伏的電位差,由鈉鉀幫浦與離子的不均勻分布共同維持。動作電位的劇本如下:
- 去極化(depolarization):當膜電位被推升到閾值,電壓門控鈉離子通道迅速開啟,鈉離子順著濃度與電位梯度湧入,把膜電位往正向推。這是一個正回饋——電位愈正,愈多鈉通道開啟——所以一旦越過閾值就「煞不住」,這就是全有全無的物理根源。
- 再極化(repolarization):鈉通道在開啟後很快去活化(inactivation),同時較慢的電壓門控鉀離子通道才打開,鉀離子外流,把膜電位拉回負值。
- 不反應期(refractory period):鈉通道去活化後需要時間才能重置,這段期間細胞無法再次放電。這個機制有兩個關鍵後果:其一,它替放電頻率設了上限(解釋了頻率編碼為何有天花板);其二,它確保動作電位只能往前傳,不會在軸突上往回倒灌——因為剛經過的那一段還在不反應期。
理解了通道,就能理解很多藥理與病理。河豚毒素(tetrodotoxin)之所以致命,是因為它專一地阻斷電壓門控鈉通道,讓動作電位無法產生;許多局部麻醉劑(如利多卡因)也是鈉通道阻斷劑,這就是為什麼打了麻藥之後痛覺訊號傳不出去。離子通道的基因突變則會造成「通道病(channelopathy)」,例如某些癲癇與心律不整。
看一個例子:閾值不是在細胞本體決定的
入門篇把「判決庭」放在軸突丘(axon hillock)。研究所層次有一個重要的精修:真正啟動動作電位的關鍵地點,是緊接在軸突丘之後、軸突最起始的一小段——軸突起始段(axon initial segment, AIS)。
為什麼是這裡而不是別處?因為 AIS 的膜上聚集了密度最高的電壓門控鈉通道,這使它成為整顆神經元中閾值最低、最容易被觸發的位置。樹突與細胞本體把所有興奮性與抑制性的輸入做時空整合,匯成一個電位,這個電位傳到 AIS,只要超過 AIS 的閾值,動作電位就在這裡點火,再往軸突傳出去。
更精彩的是,AIS 並非一成不變。近年研究發現 AIS 的長度與位置會隨神經活動而結構性地移動——當輸入長期增強,AIS 可能往遠離細胞本體的方向移動,等於提高了觸發門檻,藉此維持細胞放電的恆定(這是恆定可塑性 homeostatic plasticity 的一種形式)。也就是說,連「決策點本身」都是可塑的。下次你看到那張把軸突丘標成單一觸發點的教科書插圖,可以記得:真正的引擎室在更精確的 AIS,而且它會自己微調。
樹突的分區運算:一棵會「在地判斷」的樹
入門篇提到樹突不是被動集線盒,能產生局部的樹突棘電位(dendritic spike)。進階一點看,這牽涉到神經元的一個核心設計原則:分區化(compartmentalization)。
樹突樹不是一根均勻的天線,而是被分割成許多半獨立的小區。不同的分支可以接收不同來源的輸入、各自進行非線性運算,再把結果往細胞本體匯報。這意味著單一神經元內部就存在「多層」處理:第一層是各個樹突分支的局部整合,第二層才是細胞本體與 AIS 的全局決策。正因如此,有理論工作把具有複雜樹突的錐體細胞(pyramidal neuron)類比為一個多層人工神經網路,而不只是單一個加權求和器。
這裡涉及兩種不同的離子事件。靠近樹突遠端的分支可能產生由 NMDA 受體主導的局部去極化(NMDA spike),而某些大型錐體細胞的頂樹突(apical dendrite)上有可產生鈣離子棘波(calcium spike)的熱區。當來自感覺輸入的訊號(抵達細胞本體附近)與來自高階皮質回饋的訊號(抵達頂樹突)在時間上恰好重合,兩者會交互放大,使神經元爆發式放電(burst)。這個「兩股訊號要在不同樹突區同時到場才會強烈反應」的機制,被認為可能是知覺與回饋、底層資料與高層預期相互印證的一種生理基礎——這與優心理談的「自上而下的預期會塑造知覺」遙相呼應。
神經膠細胞:不只是包覆軸突的配角
入門篇把神經膠細胞(glia)當作製造髓鞘的「絕緣工人」,這其實低估了它們。在人腦中,膠細胞數量與神經元相當(過去「膠細胞是神經元十倍」的說法已被審慎的計數研究修正為約略相當),而且它們深度參與神經元的構造與功能。
星形膠細胞(astrocyte) 的突起會包裹突觸,形成所謂的「三方突觸(tripartite synapse)」概念:突觸前、突觸後,再加上星形膠細胞這第三方。它負責回收突觸間隙裡多餘的神經傳遞物質(例如把麩胺酸清走,避免過量造成興奮毒性),調節局部離子濃度,並支援能量供應。寡突膠細胞(oligodendrocyte) 與許旺細胞(Schwann cell) 製造髓鞘,而髓鞘的厚度與蘭氏結的間距並非隨機,是被精細調控以匹配傳導時序的——這對需要訊號「同時抵達」的神經迴路(例如聽覺定位)至關重要。微膠細胞(microglia) 則是大腦的免疫細胞,它們會主動修剪(prune)不活躍的突觸,是發育期與學習期突觸重塑的關鍵推手。
所以,當我們談「神經元的構造」,嚴格說來是在談一個由神經元與膠細胞共同構成的功能單元。把膠細胞排除在外的神經元圖,就像只畫了演員卻刪掉了整個劇組。
重點回顧
- 軸突運輸是維生命脈:蛋白質只在細胞本體合成,靠驅動蛋白(順向)與動力蛋白(逆向)沿微管往返運輸;故障會導致由末梢往內的逆死性退化。
- 全有全無源自離子通道時序:電壓門控鈉通道的正回饋造成不可逆的去極化,鉀通道與鈉通道去活化負責再極化與不反應期,後者同時設定頻率上限並保證單向傳導。
- 真正的觸發點是 AIS:軸突起始段鈉通道密度最高、閾值最低,是動作電位的點火室,且其位置會隨活動結構性地調整。
- 樹突是分區化的多層運算器:不同樹突分支各自進行 NMDA/鈣離子棘波等非線性運算,使單一神經元近似多層神經網路。
- 膠細胞是構造的共同作者:星形膠細胞調節突觸與離子環境、寡突膠/許旺細胞塑形髓鞘時序、微膠細胞修剪突觸。
深入探討(研究所視角)
進入研究所層次,「神經元的構造」會被當成一個可被定量、可被擾動、可被反向工程的系統。以下幾個方向值得延伸。
從電纜理論到區室模型。 要把樹突的分區運算寫成數學,傳統工具是 Wilfrid Rall 在 1960 年代奠基的電纜理論(cable theory),它用偏微分方程描述電位沿著樹突這條「有漏電的電纜」如何衰減與延遲。現代的計算神經科學則把神經元切成數百到數千個「區室(compartment)」,每個區室各自套用 Hodgkin–Huxley 式的通道動力學,再用電阻把相鄰區室連起來,這就是 NEURON、Brian 等模擬軟體背後的多區室模型(multi-compartment model)。理解構造的分區,正是建立這類模型的前提:你必須先知道通道分布在哪、樹突幾何長什麼樣,才能把一顆神經元變成可在電腦裡跑的方程組。
結構連接體(connectome)與顯微術的躍進。 「神經元構造」這個題目近年因技術而重生。冷凍電子顯微術(cryo-EM) 讓我們看清離子通道與受體的原子級結構,直接把「閘門怎麼開」變成可見的構象變化;血清切片電子顯微鏡 + 機器學習自動重建則讓研究者得以重建出包含每一條樹突、每一個突觸的奈米級線路圖。2024 年發表的果蠅成蟲全腦連接體(含約十四萬個神經元與數千萬個突觸)是一個里程碑,它把「構造」從單顆細胞的解剖,推進到整個迴路的精確布線——這也回過頭來驗證了分區運算與細胞型態多樣性的重要性。
與人工智慧的雙向對話。 入門篇談過人工神經元是生物神經元的簡化。進階的張力在於:當我們發現單一生物神經元的樹突就具備多層運算能力、AIS 會自我調整閾值、膠細胞還在旁邊修剪連結,這提醒我們「一個生物神經元≈一個人工權重節點」的對應其實非常粗糙。這個落差正是脈衝神經網路(spiking neural network) 與神經形態運算(neuromorphic computing) 想縮小的目標——前者讓人工單元也用離散脈衝與時序編碼,後者讓硬體在物理層次模仿離子通道的事件驅動與低功耗特性。對 Uedu「人機共學」的立場而言,認清這層差異不是要貶低 AI,而是提醒我們:在拿大腦類比深度學習時,要清楚自己簡化掉了什麼。
跨領域連結。 從優生物看,軸突運輸與細胞骨架是發育生物學、馬達蛋白生物物理的核心案例;離子通道則串起膜生理與藥理學(從河豚毒素到局部麻醉劑都在同一個分子上作用)。從優心理看,樹突的「感覺輸入×高階回饋」重合機制,是預測編碼(predictive coding)與注意力理論的可能生理對應;而膠細胞主導的突觸修剪,則為「用進廢退」這句老話提供了細胞層次的執行者。當你在不同課堂遇到「結構決定功能」這句箴言時,神經元是最有說服力的範例之一:它把運算、記憶、決策,全都寫進了自己的形狀裡。