
你昨天背的單字,今晚睡覺時去了哪裡?
從記憶痕跡(engram)到突觸長期增益(LTP),看大腦如何把經驗刻進神經元連接的強度裡。
你昨天背的單字,今晚睡覺時去了哪裡?
想像一位學習者,昨天晚上花了一小時記下二十個英文單字。今天早上,有些單字脫口而出,有些卻怎麼也想不起來。一週後,能穩定記得的或許只剩一半。這個再日常不過的經驗,背後其實是一場橫跨數百萬個神經元、歷時數小時到數天的生物化學工程。「記憶」不是被寫進某個檔案夾,而是被刻進了神經元之間連接的強度裡。
這篇文章要回答一個核心問題:當你學會一件事,你的大腦裡到底「改變」了什麼?我們會從一個被稱為「記憶痕跡」(engram)的概念出發,一路走到突觸層級的長期增益(long-term potentiation, LTP)機制,最後連到分子、系統,乃至人工智慧。
記憶痕跡:記憶有沒有「實體地址」?
早在 1904 年,德國學者 Richard Semon 就提出了 engram(記憶痕跡)這個詞,主張一段經驗會在腦中留下持久的物理痕跡。但接下來數十年,這個概念近乎只是哲學猜想。心理學家 Karl Lashley 花了三十年訓練大鼠走迷宮,再切除牠們大腦皮質的不同部位,試圖找出記憶「藏在哪裡」。結果讓他在 1950 年寫下著名的挫敗結論:他「找不到 engram」——記憶似乎廣泛分散,而非定點儲存。
Lashley 的失敗其實藏著正確的線索:記憶確實是分散式(distributed)的。真正的轉折要等到現代技術。2012 年起,利賓川光(Susumu Tonegawa,1987 年諾貝爾生醫獎得主)的團隊運用光遺傳學(optogenetics)證明:在小鼠形成恐懼記憶時被活化的那一小群海馬迴神經元,事後若用光人為地重新激活,小鼠會表現出「記得」當時情境的恐懼反應。換句話說,記憶痕跡不只存在,還可以被「點亮」。後續研究甚至能植入虛假記憶、抹除特定記憶。
這帶來一個關鍵觀念修正:engram 不是某個神經元,而是一群被特定經驗連結起來的神經元集合(engram cell ensemble)。學習,就是讓這群細胞之間的連接被選擇性地強化,使它們日後容易一起被激活。那麼,「連接被強化」具體是什麼意思?這就要進入突觸的世界。
突觸可塑性:學習發生在神經元的「握手處」
兩個神經元相接觸、傳遞訊號的地方叫突觸(synapse)。前一個神經元(突觸前)釋放神經傳導物質,後一個神經元(突觸後)接收。1949 年,心理學家 Donald Hebb 提出了一個後來被奉為圭臬的假說,常被濃縮成一句口訣:
"Cells that fire together, wire together."(一起放電的細胞,會連在一起。)
Hebb 的原始說法更精確:當細胞 A 反覆且持續地參與激發細胞 B,兩者之間的連接效率就會提升。這就是「突觸可塑性」(synaptic plasticity)的核心——突觸的強弱不是固定的,而是會根據使用經驗動態改變。學習與記憶的生物基礎,正是這種改變。
但 Hebb 的假說在 1949 年仍只是理論。直到 1973 年,Terje Lømo 與 Tim Bliss 在兔子海馬迴上做出了關鍵實驗,第一次直接觀察到突觸真的能被「強化並維持」。這個現象,就是 LTP。
LTP:把 Hebb 假說變成可測量的現象
LTP(long-term potentiation,長期增益)指的是:對一條神經通路施加短暫但高頻的電刺激後,該突觸的傳遞效率會顯著增強,而且這個增強能維持數小時、數天,甚至更久。
看一個例子:海馬迴裡的高頻刺激
讓我們具體想像 Bliss 與 Lømo 的實驗邏輯。研究者在兔子海馬迴的一條輸入通路上放電極,先測量「基準反應」:給一個標準刺激,記錄突觸後神經元產生多大的電位。
接著,他們對這條通路施加一陣高頻刺激(例如 100 Hz、持續一秒的強烈轟炸,稱為 tetanus)。
之後再用原本的標準刺激去測——驚人之處在於:同樣強度的輸入,現在卻引發了明顯更大的突觸後反應。這個「變大」可以持續很久。這就是突觸「記住了」剛剛被密集使用過。LTP 之所以被視為記憶的細胞模型,正因為它具備記憶該有的特質:
- 快速形成:幾秒的刺激就能誘發。
- 持久:可維持遠超過刺激本身的時間。
- 輸入專一性(input specificity):只有被刺激的那條通路變強,旁邊沒被刺激的不變。
- 協同性與關聯性:弱輸入若與強輸入同時發生,也能搭上順風車被強化——這正對應了「關聯學習」。
分子機制:NMDA 受體這個「巧合偵測器」
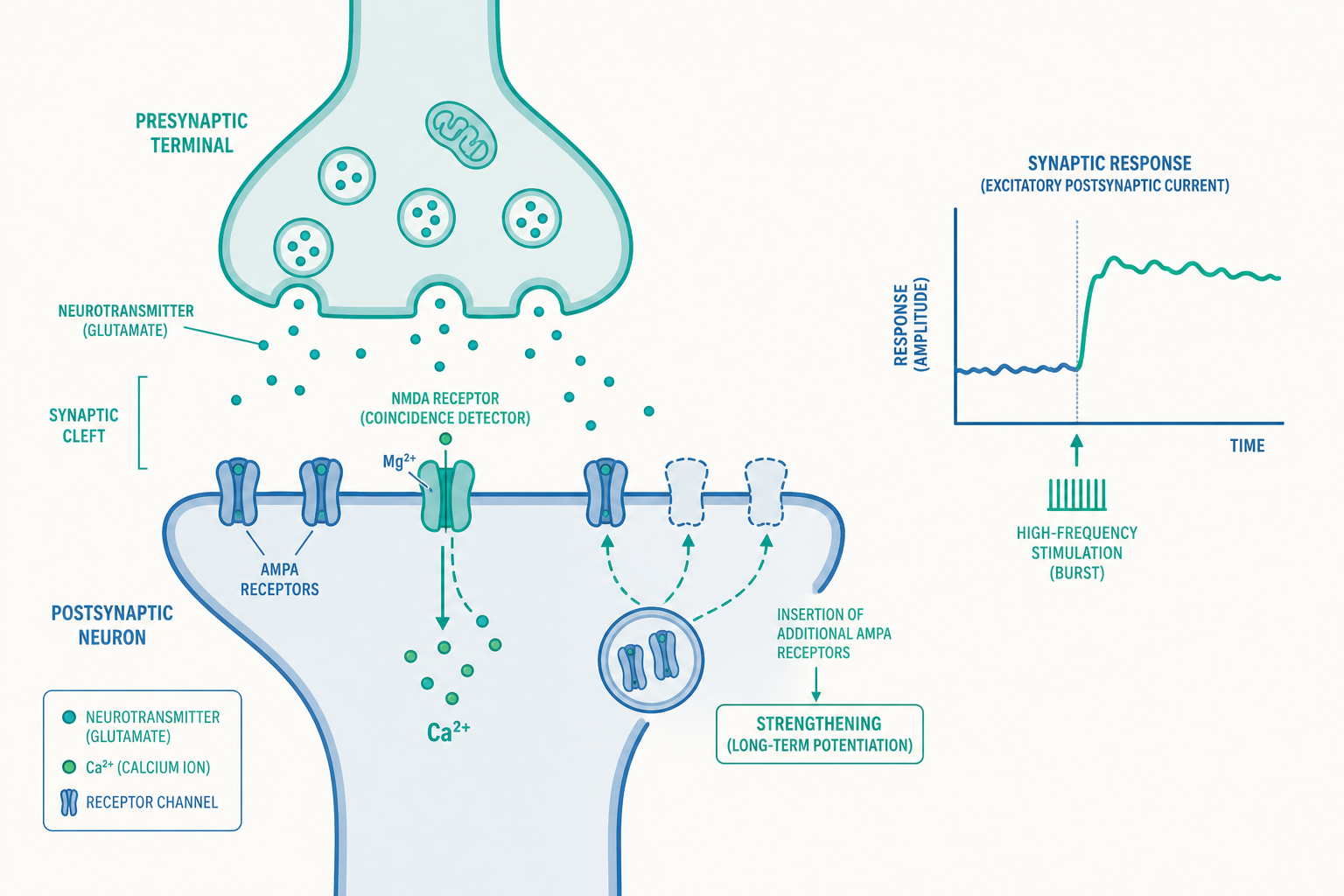
LTP 最經典的形式發生在海馬迴 CA1 區,核心角色是兩種接收麩胺酸(glutamate,主要興奮性神經傳導物質)的受體:AMPA 受體與 NMDA 受體。理解它們的分工,是理解 LTP 的關鍵。
平常傳遞訊號時,麩胺酸打開 AMPA 受體,讓鈉離子流入,產生一般的興奮。但 NMDA 受體很特別:它的通道平時被一個鎂離子(Mg²⁺)像塞子一樣堵住。要打開它,必須同時滿足兩個條件:
- 突觸前釋放麩胺酸(代表「上游有訊號來了」)。
- 突觸後膜已經被去極化(depolarized,代表「下游也正在強烈活動」)——去極化才能把鎂離子塞子推開。
因此 NMDA 受體被稱為「巧合偵測器」(coincidence detector):唯有當「前」與「後」幾乎同時活躍,它才放行。這在分子層級精準實現了 Hebb 的「一起放電」原則。
一旦 NMDA 受體開啟,鈣離子(Ca²⁺)大量湧入突觸後神經元。鈣離子是關鍵的第二信使,會啟動一連串酵素反應,其中最重要的是活化 CaMKII(一種鈣/鈣調素依賴性蛋白激酶)。接下來發生兩件改變突觸強度的事:
- 更多 AMPA 受體被插入突觸後膜:受體變多,同樣的麩胺酸就能引發更大反應——突觸「聽得更清楚」了。這是早期 LTP(early-LTP)的主要機制。
- 既有 AMPA 受體被磷酸化:傳導效率提升。
動手試試:用一個比喻自我檢查
試著向自己(或同學)解釋這個比喻,看能否說得通:把突觸想成一道門,AMPA 受體是門上現有的把手數量,NMDA 受體則是一個「需要兩把鑰匙同時轉動」才會啟動的升級開關。當上游與下游同時活躍(兩把鑰匙都轉),升級開關啟動,工人(鈣離子訊號)就跑來門上加裝更多把手。下次再來開門,就更輕鬆了。
如果你能流暢地說出「為什麼一定要兩把鑰匙」(答案:避免突觸隨便就被強化,唯有真正相關的活動才值得記住),代表你已抓住 NMDA 受體巧合偵測的精髓。
從早期 LTP 到晚期 LTP:為什麼記憶需要時間「固化」
回到開頭的單字問題。為什麼有些記憶睡一覺就鞏固了,有些卻流失?這牽涉到 LTP 的兩個階段。
早期 LTP(E-LTP) 只靠現有蛋白質的修飾(如插入既有受體、磷酸化),維持約一到三小時,不需要製造新蛋白。它對應的是短期到中期記憶。
晚期 LTP(L-LTP) 才是長期記憶的基礎。當刺激夠強或重複夠多次,鈣訊號會啟動 CREB(cAMP 反應元件結合蛋白)這類轉錄因子,開啟基因表現、合成新的蛋白質,甚至長出新的樹突棘(dendritic spine)、建立新的突觸連接。這是一種結構性的改變,能維持數天到數月。
這解釋了幾個重要的學習現象:
- 為什麼「分散練習」勝過「臨時抱佛腳」:間隔重複(spaced repetition)讓每次複習都重新觸發鞏固機制,更有機會推進到 L-LTP 的蛋白質合成階段。一次塞滿則多停留在 E-LTP,容易流失。
- 為什麼睡眠對記憶如此關鍵:睡眠(尤其慢波睡眠)期間,海馬迴會「重播」白天的神經活動序列,協助把記憶逐步轉移、固化到大腦皮質——這個過程稱為系統固化(systems consolidation)。
LTD 與雙向可塑性:記憶不只靠「加強」
如果突觸只會增強,大腦很快就會飽和。事實上,存在一個與 LTP 相反的機制:LTD(long-term depression,長期抑制)。當突觸受到低頻、微弱而持續的刺激,鈣離子是「少量緩慢」湧入時,反而會啟動另一套酵素(磷酸酶),讓 AMPA 受體被移除、突觸變弱。
LTP 與 LTD 構成雙向可塑性,讓大腦既能強化重要連結、又能修剪無用連結。這對學習至關重要:遺忘並非單純的失敗,有時是主動的、有功能的修整,讓真正重要的訊號從雜訊中浮現。
重點回顧
- 記憶痕跡(engram)是分散式的細胞集合:學習會把一群神經元連結起來,使它們日後容易共同被激活;光遺傳學已證實這群細胞可被人為「點亮」。
- 學習的物理基礎是突觸可塑性:突觸強度會依使用經驗改變,呼應 Hebb「一起放電的細胞會連在一起」的假說。
- LTP 是記憶的細胞模型:高頻刺激讓突觸傳遞長期增強,具備快速、持久、輸入專一性等特質。
- NMDA 受體是巧合偵測器:唯有突觸前釋放麩胺酸、且突觸後同時去極化,鈣離子才湧入並啟動強化——這在分子層級實現了 Hebb 原則。
- 長期記憶需要新蛋白質合成:晚期 LTP 透過 CREB 等轉錄因子改變基因表現、建立新突觸結構,這正是分散練習與睡眠有助記憶的根本原因。
深入探討(研究所視角)
突觸標記與捕獲(synaptic tagging and capture, STC)。 一個長期未解的難題是:L-LTP 需要新合成的蛋白質,但蛋白質在細胞本體製造後,如何「知道」要送到剛被活化的那一個突觸,而非整顆神經元上千個突觸?Frey 與 Morris(1997)提出的 STC 假說給了優雅解答:被強烈活化的突觸會留下一個短暫的「標記」(tag),而新合成的可塑性相關蛋白(plasticity-related proteins, PRPs)則在細胞內擴散,被任何帶標記的突觸「捕獲」並利用。這個模型解釋了行為上的「行為標記」現象——一個本來只夠形成短期記憶的弱學習,若在時間窗內伴隨另一個強烈經驗(提供 PRPs),竟能轉為長期記憶。這對教學設計極具啟發:弱、易忘的學習內容若與情緒強烈或高度投入的經驗鄰近發生,可能被一併固化。
工程化的 engram 操弄與記憶的可塑邊界。 Tonegawa 實驗室與 Sheena Josselyn、Paul Frankland 等人的後續工作,已能標記、再激活、抑制甚至「重寫」engram。Ramirez 與 Liu 等人(2013)植入虛假恐懼記憶;也有研究顯示活化正向記憶的 engram 可緩解小鼠的憂鬱樣行為。這引出一個深刻的理論辯論:記憶究竟「儲存在突觸權重」,還是部分「儲存在細胞固有興奮性與核內表觀遺傳狀態」?近年表觀遺傳學(DNA 甲基化、組蛋白修飾)研究指出,記憶的某些成分可能編碼在比突觸更穩定的層級,這對「突觸即記憶」的正統觀點提出了補充與挑戰。
跨領域連結——通往人工智慧。 Hebbian learning 是人工神經網路的歷史源頭:「依共同活化調整連接權重」直接啟發了早期學習規則,而現代深度學習的反向傳播(backpropagation)雖在生物上不直接對應,卻共享「以經驗調整突觸/權重」的核心精神。值得學習者深思的對照是「災難性遺忘」(catastrophic forgetting):人工神經網路學新任務時會抹除舊任務的權重,而生物大腦透過互補學習系統(complementary learning systems)——海馬迴快速學習、皮質緩慢固化——以及突觸鞏固機制(如 elastic weight consolidation 等演算法所模仿的)來避免遺忘。這條從 NMDA 受體到人工智慧的線索,正是 Educational Omics 中 Cognomics(認知歷程)與當代 AI 對話的交會點。
回到課堂的整合視角。 把分子(NMDA/鈣/CREB)、突觸(LTP/LTD)、系統(海馬迴—皮質固化、睡眠重播)與行為(間隔練習、提取練習)串成一條因果鏈,學習者就能理解:許多被實證支持的學習策略——分散練習、檢索練習(retrieval practice)、充足睡眠——並非武斷的讀書技巧,而是順著突觸可塑性與記憶固化的生物時程而設計。當你理解了大腦如何把經驗刻進連接強度裡,你也就握有了更聰明地與自己大腦合作的鑰匙。