
一個念頭,要跨過多少道「閘門」才到得了你的手指?
從樹突、細胞本體、軸突到突觸,拆解神經元如何以「接收—整合—輸出—傳遞」完成一次思考與反應。
一個念頭,要跨過多少道「閘門」才到得了你的手指?
當你看到這行字、決定點頭的那一瞬間,你的大腦裡有數以億計的細胞在彼此低語。它們不是用喊的,而是用一種精巧到令人讚嘆的方式傳遞訊息:先在細胞內部以電流奔跑,到了細胞與細胞之間的縫隙,又改用化學分子「擺渡」過去。這個過程的主角,就是神經元(neuron,又稱神經細胞)。
一個神經元看起來像什麼?如果你把它畫下來,它既不像皮膚細胞那樣方方正正,也不像紅血球那樣圓滾滾。它有一叢像樹枝般往外散開的觸手、一個鼓鼓的中心,再加上一條細長的「電纜」。這三個部位,分別對應著神經系統運作的三個基本動作:接收、整合與輸出。理解它們的構造,就是理解「思考」這件事在物理層次上到底發生了什麼。
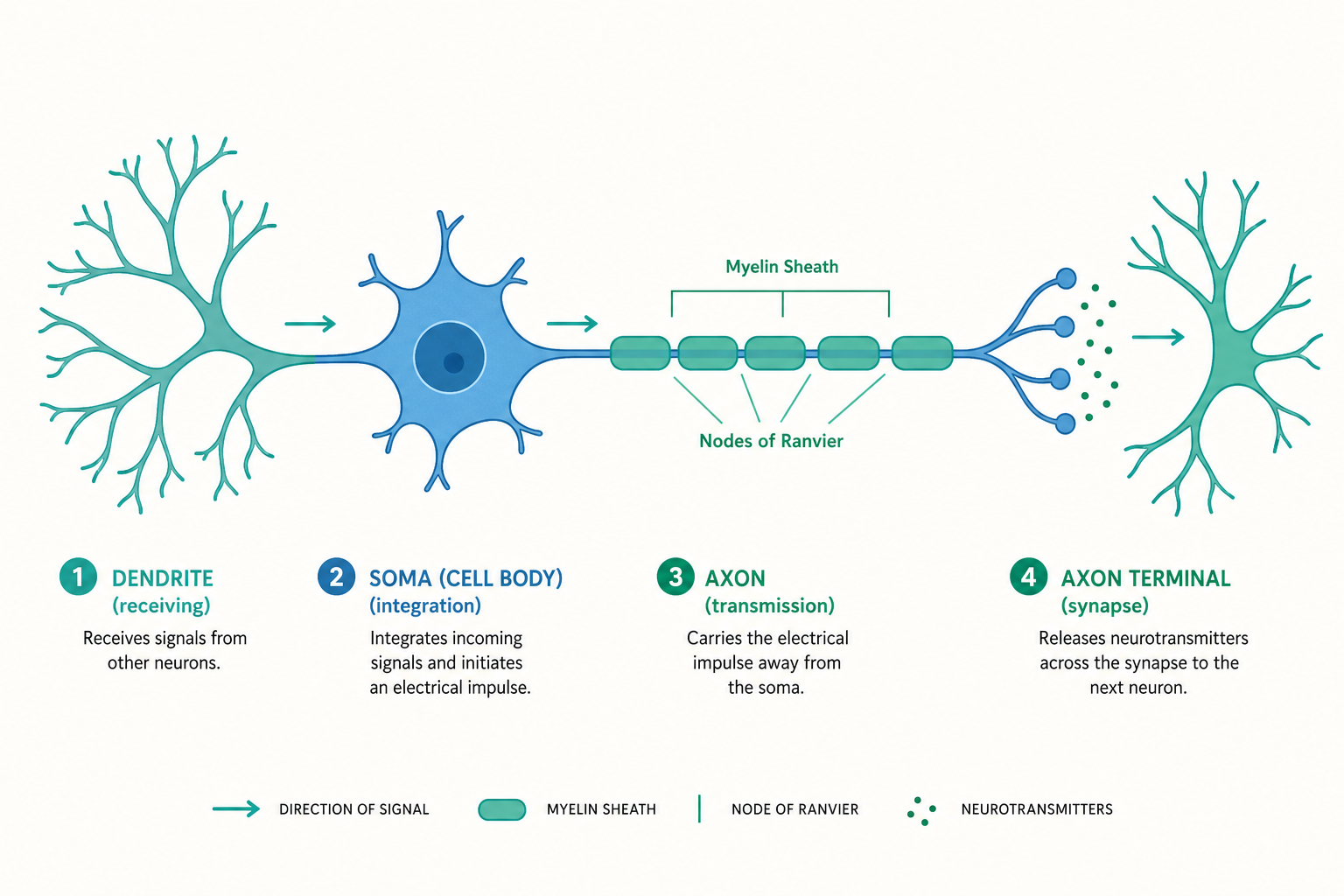
樹突(Dendrite):訊息的入口
樹突是神經元上那一叢往外分岔的突起,名字來自希臘文 déndron(樹),因為它的分支型態確實神似一棵樹的枝枒。樹突的工作是接收來自其他神經元的訊息。
你可以把樹突想成一個細胞的「天線陣列」。一個神經元的樹突表面可能佈滿了成千上萬個接收點,每個接收點都準備好迎接前一個神經元送來的化學訊號。樹突的分支愈繁密、表面積愈大,這個神經元能同時聽取的「對話」就愈多。大腦皮質中的錐體細胞(pyramidal neuron)就有極為發達的樹突樹(dendritic tree),這也是它能進行複雜整合的結構基礎。
在許多神經元的樹突上,還有一種更微小的構造叫樹突棘(dendritic spine),是一個個從樹突表面突出的小蘑菇狀構造。大多數興奮性突觸就落在這些棘上。值得一提的是,樹突棘並非固定不變——它們會隨著學習與經驗改變形狀、數量,甚至生成或消失。這種結構上的可塑性,正是「學習會改變大腦」這句話的微觀證據之一。
需要破除一個常見迷思:樹突並不是被動的「集線盒」。長久以來教科書傾向把樹突描繪成只負責收集訊號、再老老實實送到細胞本體的電線。但近數十年的研究顯示,樹突本身就具有運算能力,某些分支能產生局部的電位事件(dendritic spike),相當於在訊號抵達細胞本體之前,先做了一層初步的「在地判斷」。換句話說,運算不是只發生在細胞中央,而是分散在整棵樹突樹上。
細胞本體(Soma):整合與決策中心
樹突收來的訊號最終匯聚到細胞本體(soma,又稱 cell body)。這裡是神經元的代謝中樞,包含細胞核以及製造蛋白質、維持細胞存活所需的各種胞器。從生命維持的角度看,細胞本體就是這個細胞的「總部」。
但它更關鍵的角色是整合與決策。樹突送進來的訊號有的是興奮性(讓細胞更傾向放電)、有的是抑制性(讓細胞更不傾向放電)。這些訊號在空間上(來自不同樹突)與時間上(在不同時刻抵達)疊加在一起,這個過程稱為整合(summation)。
細胞本體與軸突交界處有一個關鍵地點,叫軸突丘(axon hillock)。這裡可以視為神經元的「判決庭」:當疊加後的電位變化超過某個臨界值——也就是閾值(threshold)——神經元就會「決定」發射一個動作電位(action potential)。這是一個全有全無(all-or-none)的事件:要嘛產生一個完整的動作電位,要嘛什麼都不發生,沒有「發射一半」這種事。訊號的強弱不是靠單一脈衝的大小來編碼,而是靠放電的頻率——刺激愈強,神經元放電愈密集。
軸突(Axon):訊號的高速公路
一旦軸突丘決定放電,動作電位就沿著軸突(axon)這條細長的纖維向外傳遞。軸突是神經元的輸出通道,把訊息送往下一個目的地。軸突的長度差異極大:腦中某些中間神經元的軸突可能只有不到一毫米,而從脊髓延伸到腳趾的運動神經元,其軸突在成人身上可以超過一公尺——是單一細胞跨越的驚人距離。
許多軸突外面包覆著一層髓鞘(myelin sheath),這是由神經膠細胞(在中樞神經系統是寡突膠細胞 oligodendrocyte,在周邊神經系統是許旺細胞 Schwann cell)纏繞而成的脂質絕緣層。髓鞘並非連續無縫,而是一節一節的,節與節之間的裸露縫隙稱為蘭氏結(node of Ranvier)。
髓鞘的作用是大幅加快傳導速度。動作電位不必在整條軸突上一格一格慢慢爬,而是從一個蘭氏結「跳」到下一個蘭氏結,這稱為跳躍式傳導(saltatory conduction)。有髓鞘的軸突傳導速度可達每秒上百公尺,遠快於無髓鞘軸突。這也說明了為何脫髓鞘疾病(如多發性硬化症,multiple sclerosis)會造成嚴重的神經功能障礙:絕緣層受損,訊號就傳得又慢又不可靠。
突觸(Synapse):細胞與細胞之間的擺渡
兩個神經元並不是直接「焊」在一起的。軸突末端與下一個細胞之間有一道極窄的縫隙,這個訊息傳遞的接點就稱為突觸(synapse)。突觸前神經元的軸突末梢、突觸後細胞的膜,以及兩者之間約二十奈米寬的突觸間隙(synaptic cleft),共同構成了訊息交棒的舞台。
絕大多數人類神經元之間是化學突觸(chemical synapse)。當動作電位抵達軸突末梢,會觸發鈣離子流入,促使裝著神經傳遞物質(neurotransmitter)的小囊泡與細胞膜融合,把傳遞物質釋放到突觸間隙中。這些分子飄過縫隙,與突觸後膜上的受體(receptor)結合,進而改變突觸後細胞的電位狀態。電訊號就這樣被翻譯成化學訊號,再翻譯回電訊號——這是神經系統最核心的「換手」機制。
常見的神經傳遞物質包括麩胺酸(glutamate,主要的興奮性傳遞物質)、γ-胺基丁酸(GABA,主要的抑制性傳遞物質)、多巴胺(dopamine)、血清素(serotonin)、乙醯膽鹼(acetylcholine)等。許多影響情緒、認知與行為的藥物,作用點正是在突觸——例如某些抗憂鬱藥會延長血清素停留在突觸間隙的時間。
突觸的另一個迷思值得澄清:突觸的「強度」並非固定不變。同一個突觸可以變強或變弱,這種能力叫突觸可塑性(synaptic plasticity)。1949 年,神經心理學家 Donald Hebb 提出一個影響深遠的原則,常被簡化為「一起放電的神經元會連結在一起(neurons that fire together, wire together)」。這個概念後來在長期增益(long-term potentiation, LTP)的實驗中得到生理證據——這正是學習與記憶在突觸層次的物質基礎。
看一個例子:你被熱水燙到的那 0.1 秒
假設你的手指碰到滾燙的鍋子。讓我們用神經元的構造把這 0.1 秒拆解開來:
- 接收:皮膚裡感覺神經元的末梢偵測到高溫與組織傷害,產生電位變化。
- 整合:訊號在細胞本體疊加,超過閾值。
- 輸出:動作電位沿著感覺神經元的軸突(有髓鞘、跳躍式傳導,因此飛快)衝進脊髓。
- 擺渡:在脊髓的突觸處,感覺神經元釋放神經傳遞物質,激活中間神經元,再激活運動神經元。
- 反應:運動神經元的軸突一路延伸到你手臂的肌肉,命令它收縮——你的手在你「意識到痛」之前就已經縮回來了。
這個反射弧之所以能比意識更快,正是因為它走的是脊髓的捷徑,不必先繞到大腦皮質「報備」。整個過程,剛好把樹突、細胞本體、軸突、突觸這四個構造的分工演示了一遍。
重點回顧
- 樹突(dendrite)負責接收:分支狀的突起,表面常有樹突棘,且具有局部運算能力,不只是被動集線。
- 細胞本體(soma)負責整合與決策:維持細胞生命,並在軸突丘以「全有全無」的方式決定是否發射動作電位;訊號強弱由放電頻率編碼。
- 軸突(axon)負責輸出:細長纖維,髓鞘與蘭氏結使其能進行跳躍式傳導,大幅加快速度。
- 突觸(synapse)負責跨細胞傳遞:以化學方式(神經傳遞物質)越過突觸間隙,並具有可塑性,是學習與記憶的物質基礎。
- 方向性是核心原則:訊息流向為「樹突 → 細胞本體 → 軸突 → 突觸 → 下一個神經元的樹突」,依序對應接收、整合、輸出、傳遞。
深入探討(研究所視角)
進入研究所層次後,「神經元的構造」會從靜態的解剖圖,轉變為一套動態的、可被數學描述的訊號處理系統。以下幾個方向值得延伸思考。
從結構到計算:神經元作為運算單元。 經典的 Hodgkin–Huxley 模型(1952,兩人因此獲 1963 年諾貝爾生理醫學獎)以鈉、鉀離子通道的電導變化,用微分方程精確刻畫了動作電位的產生。這套模型把「軸突膜」這個生物構造,翻譯成可計算的電路。更進一步,樹突的主動特性(dendritic spike、非線性整合)使單一神經元的運算能力遠超過早期「線性加總器」的假設——有理論工作甚至主張,一個具有複雜樹突樹的錐體細胞,其計算能力近似一個多層的人工神經網路。這對「神經元是大腦最小運算單元」的傳統觀點提出了有趣的修正。
突觸可塑性的分子機制。 LTP 的核心之一是 NMDA 受體,它同時是「電壓偵測器」與「化學偵測器」:唯有當突觸後膜已去極化、且突觸前釋放麩胺酸時,它才允許鈣離子流入,從而符合 Hebb 原則的「同時性」要求。鈣訊號接著啟動一連串下游反應(如 CaMKII 活化、AMPA 受體插入),使突觸變強。這提供了一個分子層次的解答:經驗如何「寫入」大腦。延伸閱讀可關注 spike-timing-dependent plasticity(STDP),它把可塑性的方向與前後神經元放電的精確時序綁定。
與人工智慧的對話。 今日深度學習的「人工神經元」是對生物神經元的高度簡化——加權求和再過一個非線性激活函數,其靈感正來自「樹突加總、軸突丘閾值決策」這個構造。但兩者差異巨大:生物神經元以離散的脈衝(spike)在時間軸上編碼訊息,能耗極低;人工神經元則傳遞連續數值。這個落差催生了脈衝神經網路(spiking neural network, SNN) 與神經形態運算(neuromorphic computing) 等研究領域,試圖讓晶片更貼近真實神經元的時序編碼與低功耗特性。對 Uedu 平台而言,理解這層生物—人工的對照,有助於在「人機共學」的框架下,更謹慎地看待 AI 與人腦的相似與相異。
跨領域連結。 從優生物的角度,神經元的構造是發育生物學與細胞骨架(微管沿軸突運輸物質)的精彩案例;從優心理的角度,突觸可塑性是聯結論學習理論(associative learning)的生理對應,也是理解記憶、注意力、情緒調節的基礎。當你在優心理讀到「練習能強化記憶」、在 AI 課讀到「權重會隨訓練更新」,請記得:這兩句話在神經元的突觸層次,其實指向同一件事——連結的強度,會因使用而改變。