
同一個神經元,為什麼有時一觸即發、有時打死不動?
深入通道蛋白的電壓感測、被動電纜整合、軸突起始段點火與興奮性的動態調音,看動作電位如何是一台可被精密調校的類比—數位計算機。
同一個神經元,為什麼有時一觸即發、有時打死不動?
你已經知道動作電位(action potential)是「全有全無(all-or-none)」的爆發,過了閾值(threshold)就剎不住車。但這裡藏著一個入門篇沒回答的麻煩:閾值並不是一個刻在石頭上的固定數字。
同一個神經元,在你睡眠不足、咖啡因爆表、或剛被一陣抑制性輸入轟炸過之後,它的「易燃程度」可以差很多。更弔詭的是——動作電位到底「從哪裡開始」?我們直覺以為訊號從細胞本體(soma)出發、沿著軸突傳出去,但真相是它通常誕生在一個你可能沒聽過的微小構造裡。
這篇進階文章不再重述「Na⁺ 進、K⁺ 出」的基本劇情,而是往下鑽到三個層次:通道蛋白怎麼「感覺」電壓、訊號在點火之前如何被被動電纜過濾與整合、以及神經元如何動態調節自己的興奮性(excitability)。理解這些,你會發現動作電位遠不只是一個開關,而是一台可被精密調校的類比—數位混合計算機。
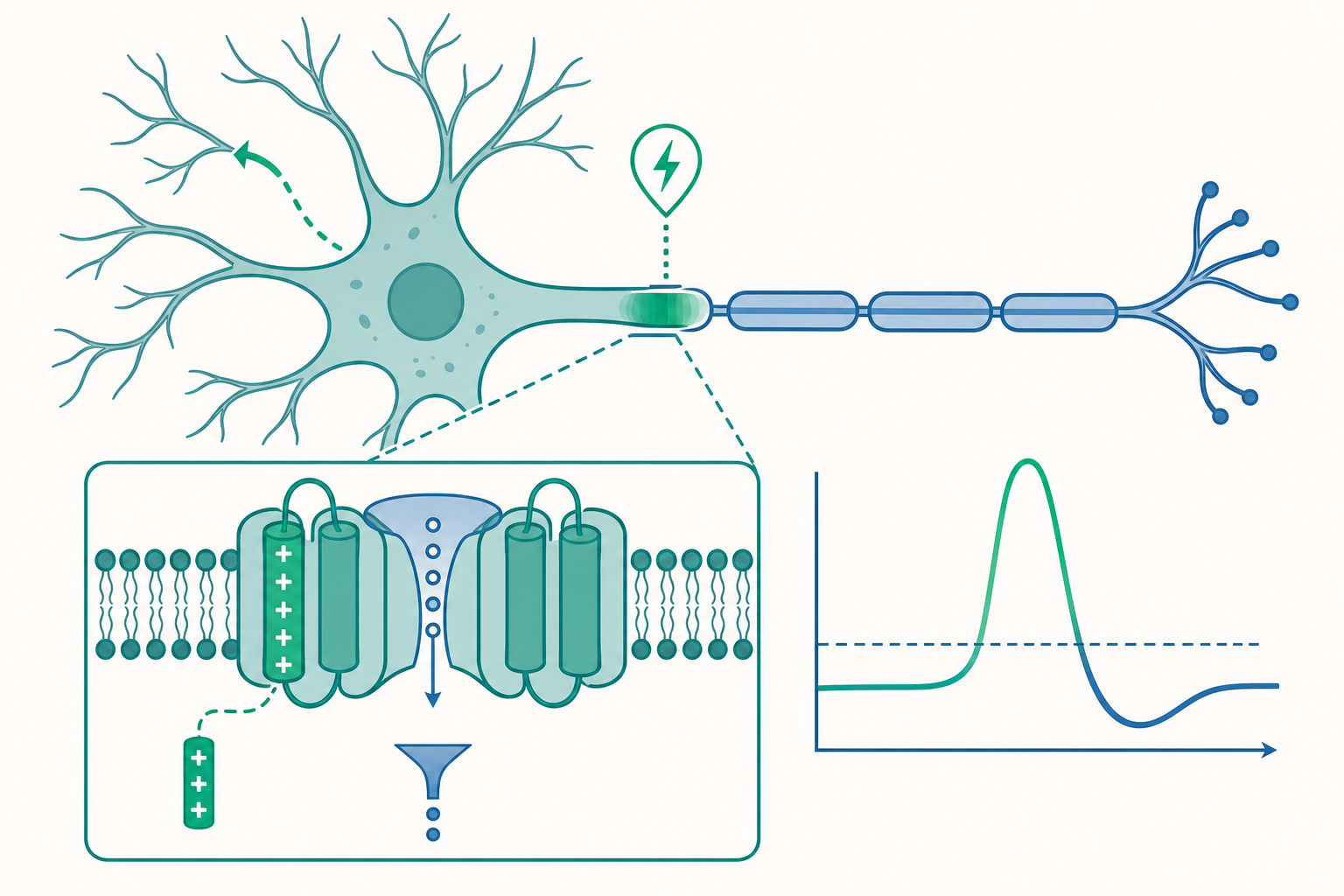
通道怎麼「看見」電壓:S4 電壓感測器與選擇性濾器
入門篇說「電壓門控鈉通道(voltage-gated Na⁺ channel)對膜電位很敏感」。但一個蛋白質憑什麼能感覺到膜內外的電場?這不是比喻,而是有具體結構基礎的。
電壓門控離子通道(如 Naᵥ、Kᵥ)的跨膜片段中,有一段稱為 S4 螺旋(S4 helix)的結構,上面每隔三個胺基酸就帶一個正電荷(多為精胺酸 arginine)。當膜去極化、膜內變正時,這串正電荷受電場推動而往細胞外側移動約 1 奈米。這個物理位移透過連桿(linker)牽動通道的活化閘門(activation gate)打開。換句話說,通道靠的是「帶電零件在電場中被推動」——這個移動本身會產生一個極微小、可被精密儀器測到的閘門電流(gating current),是 S4 真的在動的直接證據。
另一個精巧之處是選擇性濾器(selectivity filter)。鈉通道為什麼只讓 Na⁺ 通過、卻擋住更小的 K⁺?關鍵不在「篩孔大小」(直覺上小離子更該通過才對),而在於濾器內壁的羰基氧原子排列,能模擬出離子在水中的水合殼層(hydration shell)。Na⁺ 的水合幾何與濾器吻合、能量上划算;K⁺ 則「卡」得不夠服貼。鉀通道的濾器則反過來,用著名的 TVGYG 序列精準容納去水合的 K⁺。這種「不是看大小,而是看能量匹配」的選擇機制,由 Roderick MacKinnon 以 X 光晶體結構解開,獲 2003 年諾貝爾化學獎。
為什麼這對學生重要?因為它把「動作電位」從一個生理學現象,下接到了結構生物學與蛋白質物理。一個點突變改變濾器或 S4 上的一個胺基酸,就可能讓通道的電壓敏感性或選擇性失常——這正是許多離子通道病(channelopathy)的分子根源,連結到優生物的學習地圖。
點火之前:被動電纜如何「過濾」訊號
一個常見迷思是:突觸輸入一進來,神經元立刻判斷「過閾值就點火」。但在點火之前,訊號得先穿過樹突(dendrite)與細胞本體這段「不會放大、只會衰減」的被動電纜(passive cable)。這段旅程決定了什麼樣的輸入才有資格觸發動作電位。
神經元的膜可以用兩個被動參數刻畫:
- 膜電阻 Rₘ 與膜電容 Cₘ:兩者相乘得到膜時間常數(membrane time constant)τ = Rₘ × Cₘ,典型約 10–20 毫秒。τ 決定膜電位對輸入「反應有多慢、記憶有多長」。τ 大,一個突觸電位衰減得慢,就更容易與後續輸入時間總和(temporal summation)。
- 空間常數(length constant)λ:一個局部去極化沿著被動樹突傳播時,會按指數衰減,傳到 λ 距離時只剩約 37%(1/e)。λ 越大,遠處樹突的輸入越能「活著」抵達點火區。
這兩個常數合起來,使神經元成為一個時空整合器(spatiotemporal integrator):它不是被動接收,而是依據輸入的時間先後與空間位置,決定要不要把膜電位推過閾值。被動電纜理論(cable theory)由 Wilfrid Rall 在 1960 年代奠基,把神經元的複雜樹突幾何化為可計算的電路,是計算神經科學的另一塊基石。
這也解釋了一件事:遠端樹突的突觸,影響力通常比近端的小——因為訊號傳到胞體時已被衰減。但神經元演化出許多補償手段(如樹突上的主動通道),讓遠端輸入不至於完全被埋沒,這引出下一節的主角。
看一個例子:動作電位其實誕生在「軸丘」
如果動作電位的點火閾值在整個神經元上一致,那它應該在膜電位最先被推高的地方爆發——可能是某個被強烈刺激的樹突。但實驗(如膜片鉗與電壓敏感染料成像)反覆顯示:在多數神經元中,動作電位幾乎總是先在一個叫 軸丘/軸突起始段(axon initial segment, AIS)的地方誕生,再從那裡往兩個方向傳。
原因是:AIS 上的電壓門控鈉通道密度遠高於胞體與樹突(部分研究估計可達數十倍),由 ankyrin-G 等支架蛋白把通道錨定在此。高密度意味著這裡的局部閾值最低,於是它成了整個神經元的「決策點」——所有樹突整合後的電位匯流到此,只有當 AIS 被推過閾值,神經元才算真正「發言」。
更精彩的是,AIS 的位置與長度並非固定。研究發現它會隨活動經驗緩慢移動或變長(structural plasticity of the AIS):長期高活動可能讓 AIS 往軸突遠端移動以降低興奮性,反之亦然。這是一種把「我最近被激發得多不多」寫進結構、再回頭調節自身興奮性的恆定調節(homeostasis)。一個 1 毫秒尺度的快速事件,竟與數小時到數天尺度的慢速結構變化耦合在一起——這正是神經系統跨時間尺度運作的縮影。
反向傳播:動作電位也會「往回跑」進樹突
入門篇強調動作電位靠不反應期(refractory period)確保單向前進,這在軸突方向上沒錯。但故事有個常被忽略的續集:在 AIS 點火後,動作電位除了沿軸突傳出去,也會逆向侵入樹突,這稱為反向傳播動作電位(back-propagating action potential, bAP)。
bAP 的意義在於它是一個「我剛剛發放了」的回傳訊號。當 bAP 抵達某個樹突分支,正好碰上該分支此刻有突觸輸入,這個時間上的巧合會被偵測到——bAP 帶來的去極化與輸入造成的局部電位疊加,可觸發鈣離子(Ca²⁺)大量內流。Ca²⁺ 訊號正是觸發突觸可塑性(synaptic plasticity)的關鍵第二信使。
這直接連到學習理論中著名的 spike-timing-dependent plasticity(STDP,脈衝時序依賴可塑性):若突觸輸入在動作電位之前幾毫秒到達(輸入「預測」了發放),該突觸傾向增強;若在發放之後才到,則傾向減弱。bAP 提供的正是這個「之前/之後」的物理判讀工具。換句話說,動作電位不只是輸出訊號,它同時是一個寫回樹突、用來標記哪些輸入該被強化的教學訊號——這把細胞層次的電生理,直接接到了優心理所關心的學習與記憶機制。
興奮性不是常數:神經元如何被「調音」
回到開頭的問題:為什麼同一個神經元時而易燃、時而冷淡?因為神經元的興奮性是一個被多重因素持續調節的變數,而非固定設定。
- 神經調節物質(neuromodulators):乙醯膽鹼、血清素、多巴胺、正腎上腺素等不像快速傳導物質那樣直接開關離子通道,而是透過 G 蛋白偶聯受體(GPCR)與細胞內訊號級聯,改變某些 K⁺ 通道的開放程度,進而調整閾值、不反應期與發放模式。例如,乙醯膽鹼可關閉某類「漏鉀」通道,讓神經元更易被持續激發——這與注意力與清醒狀態的調控密切相關。
- 發放後適應(spike-frequency adaptation):許多神經元在持續輸入下,發放頻率會逐漸下降。機制之一是 鈣活化鉀通道(Ca²⁺-activated K⁺ channel):每次發放都讓一點 Ca²⁺ 進來,累積的 Ca²⁺ 打開這類 K⁺ 通道、產生後超極化(afterhyperpolarization, AHP),使後續發放更難。這是一種內建的「自我節流」。
- 恆定性可塑性(homeostatic plasticity):在更長時間尺度上,神經元會調整離子通道的表現量,把自己的平均發放率拉回某個目標範圍,避免因學習導致的突觸增強而陷入失控或沉默。
把這三層放在一起,動作電位的閾值就像音響上一個會自動調整的旋鈕:快(毫秒)的不反應期、中(秒)的發放適應、慢(小時到天)的恆定調節,共同決定此刻這個神經元有多容易「說話」。
動手試試:用能量帳本看 spike 的代價
動作電位不是免費的。每發放一次,內流的 Na⁺ 必須由鈉鉀幫浦(Na⁺/K⁺-ATPase)打回去,而幫浦每搬運離子就要消耗 ATP。試著粗估一下這筆帳:
- 大腦只佔體重約 2%,卻消耗全身約 20% 的靜息能量——這是教科書級的事實,不是估算。
- 其中很大一部分花在維持離子梯度與支應動作電位後的離子復原。研究(如 Attwell 與 Laughlin, 2001 的能量預算分析)估計,突觸傳遞與動作電位是皮質能量消耗的主要去處。
- 推論:演化會傾向讓編碼節能。這正是「稀疏編碼(sparse coding)」假說的能量論據之一——若每個刺激只讓少數神經元發放,就能在維持資訊量的同時省下大量 ATP。
試著反過來想:如果某種疾病(如缺血、粒線體功能障礙)讓 ATP 供應不足,會先發生什麼?答案是離子梯度無法維持、膜電位失控去極化、通道與幫浦失靈——這正是中風核心區神經元死亡的電生理鏈條。一個看似抽象的「能量帳本」,直接連到臨床上分秒必爭的救治窗口。
重點回顧
- 通道靠帶電的 S4 螺旋「看見」電壓:去極化推動 S4 上的正電荷位移約 1 奈米,開啟閘門;選擇性濾器則靠能量匹配(而非孔徑大小)區分 Na⁺ 與 K⁺。
- 點火之前先過被動電纜:膜時間常數 τ 與空間常數 λ 使神經元成為時空整合器,決定哪些輸入有資格觸發動作電位。
- 動作電位誕生於軸突起始段(AIS),因該處鈉通道密度最高、閾值最低;AIS 位置還會隨活動經驗緩慢重塑,達成興奮性恆定。
- 反向傳播動作電位(bAP)把「我剛發放了」的訊號送回樹突,提供 STDP 所需的時序判讀,是學習的細胞層次橋樑。
- 興奮性是動態變數:神經調節物質、發放適應、恆定可塑性在毫秒到天的尺度上共同「調音」;而每次發放都有實打實的 ATP 代價。
深入探討(研究所視角)
從「點」到「分布」:通道的隨機性與通道雜訊。 入門模型把離子電導當成平滑連續的量,但真實通道是離散且隨機開關的。單一通道的開關是機率事件,在通道數目少(如細軸突或小的興奮區)時,這種通道雜訊(channel noise)會明顯影響閾值與發放時機,甚至可自發觸發動作電位。這把確定性的 Hodgkin–Huxley 框架推向隨機(stochastic)建模——以化學主方程式(master equation)或 Langevin 近似描述通道狀態的機率演化。對研究者而言,這意味著「神經元是否發放」在微觀尺度上帶有不可化約的隨機性,而這種隨機性未必是缺陷:在某些情況下,適度雜訊反而能透過隨機共振(stochastic resonance)提升對微弱訊號的偵測——一個橫跨物理與神經科學的迷人現象。
軸突不是被動電線:軸突計算(axonal computation)。 入門篇把軸突描繪為忠實傳遞數位 spike 的通道,但前沿研究顯示軸突本身可能參與運算。閾下的膜電位變化能沿軸突傳播一定距離,調節到達末梢時的突觸釋放量(即類比—數位混合訊號,analog-digital signalling);軸突的分支點(branch point)幾何還可能造成傳導失敗(propagation failure)或分支間的差異傳導,使同一個動作電位在不同分支產生不同效果。這顛覆了「軸突只是一條全有全無的電線」的簡化圖像,暗示單一神經元的計算能力被長期低估。
跨領域連結:生物的精巧 vs. 人工網路的抽象。 把本文的機制與人工智慧並置,反差很有啟發性。標準人工神經元把「整合輸入 → 過閾值 → 輸出一個數」壓縮成一次靜態的加權求和加激活函數,丟掉了時間動態、樹突的非線性整合、AIS 的空間結構、與 bAP 的回傳教學訊號。近年的樹突計算(dendritic computation)研究指出,單一生物神經元因樹突上的主動通道,可能等價於一個小型多層網路,本身就能解決異或(XOR)這類非線性可分問題——這是單層感知器(perceptron)做不到的。脈衝神經網路(SNN)與神經形態運算(neuromorphic computing)正試圖把時間維度與事件驅動的稀疏發放找回來,但要提醒:這些仍是選擇性借用生物原理,而非複製神經元。理解「生物多保留了什麼、AI 抽象掉了什麼」,本身就是跨領域研究最有價值的提問。
一個仍在演進的開放問題。 動作電位是教科書寫了七十年的「定案」機制,但它的細節仍在被改寫。AIS 的精確結構與可塑性、軸突的類比成分、通道雜訊的功能角色、乃至膠細胞(glia)如何透過調節細胞外鉀濃度與髓鞘厚度間接影響傳導——這些都還是活躍的研究前沿。對準備進入研究所的你,這帶來一個重要的態度:經典不等於完結。能在一個看似已被講透的機制裡,找到尚未被回答的縫隙,正是研究者最核心的能力。