
當你伸手去接一只滑落的咖啡杯
運動系統如何在毫秒之間,把抽象的「意圖」轉換成精準協調的動作。
當你伸手去接一只滑落的咖啡杯
想像你正坐在書桌前,手肘不小心碰到杯緣,杯子開始往桌邊傾倒。在你「意識到」之前,你的手已經張開、加速、轉向,在杯子墜地前一瞬間穩穩接住。整個動作不到三百毫秒,比你能清楚說出「危險」這個念頭還要快。
這個看似輕鬆的反應,其實動用了大腦與脊髓裡一整套龐大而精密的協調系統。它要在毫秒之間決定:哪些肌肉該收縮、收縮多大力、依什麼順序、又要如何根據視覺即時修正。我們把這套負責「規劃動作」與「執行動作」的神經機制,統稱為運動系統(motor system)。
運動系統是神經科學中少數能讓我們「看見」大腦輸出的窗口。感覺輸入很多時候難以驗證,但動作是公開可觀察的——一隻顫抖的手、一個遲疑的步伐,往往就是神經迴路狀態的外顯證據。理解運動系統,等於理解大腦如何把「意圖」轉換成「世界裡的物理變化」。
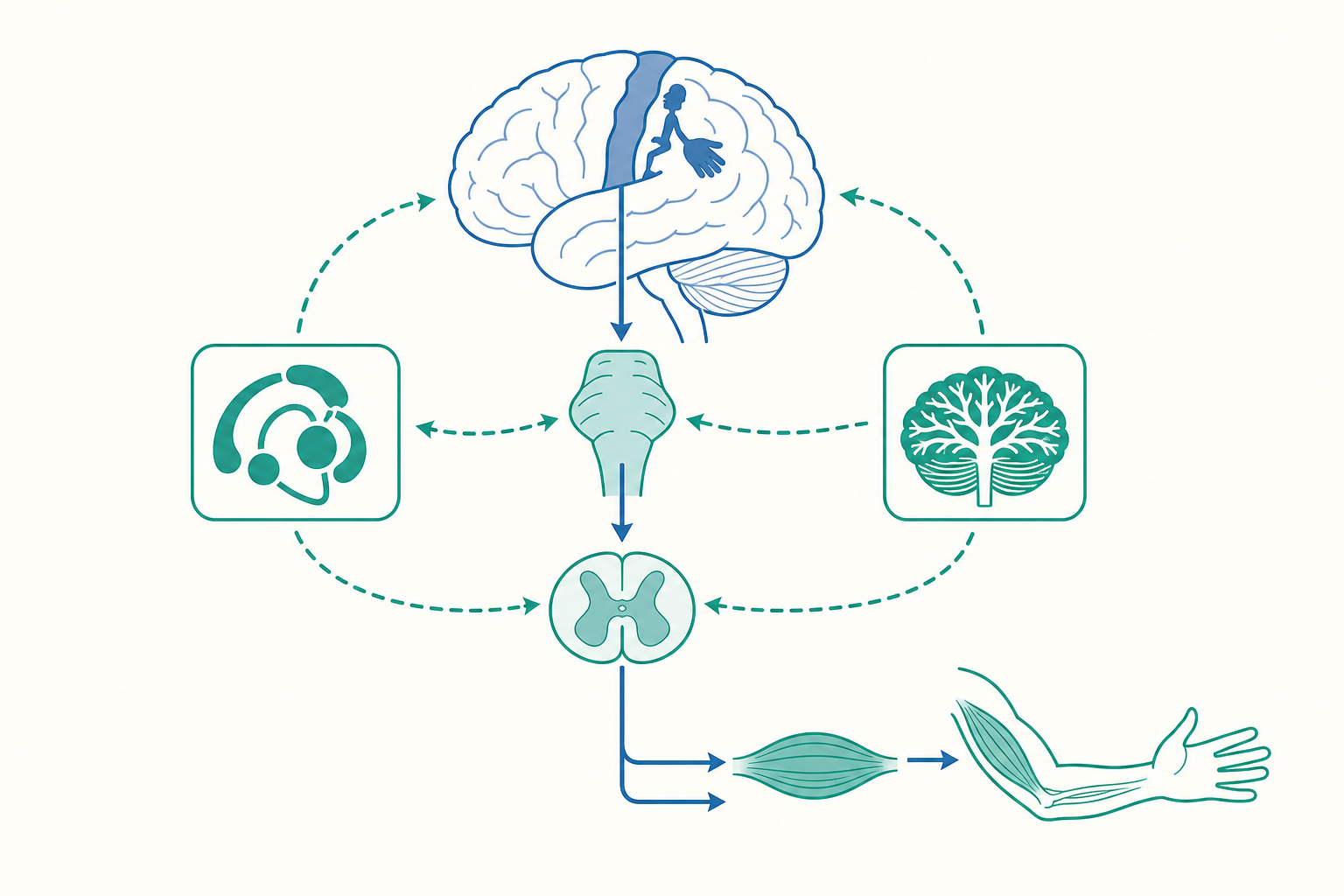
從意圖到動作:一條多層級的指揮鏈
人類的動作控制不是單一中樞下命令,而是一個階層式(hierarchical)且平行(parallel)的分工系統。我們可以把它粗略分成三個層級。
最高層:規劃與決策。 前額葉皮質(prefrontal cortex)與後頂葉皮質(posterior parietal cortex)負責整合目標、情境與感覺資訊,形成「我要做什麼」的抽象意圖。例如「接住杯子」這個目標本身,並不指定任何特定肌肉。
中間層:策略轉換為運動程式。 前運動皮質(premotor cortex)與輔助運動區(supplementary motor area, SMA)把抽象目標轉成具體的動作序列與時序安排。前運動皮質偏向處理由外在線索引導的動作(看到杯子滑落而反應),輔助運動區則偏向內在發動、需要記憶序列的動作(憑記憶按出一串密碼)。
最低層:執行。 初級運動皮質(primary motor cortex, M1)位於中央前回(precentral gyrus),是大腦皮質下達肌肉指令的主要出口。M1 的神經元軸突組成皮質脊髓束(corticospinal tract),一路下行到脊髓,與運動神經元連結,最終驅動肌肉。
這個階層並非嚴格的單向命令。各層級之間有大量回授迴路,且基底核(basal ganglia)與小腦(cerebellum)這兩個皮質下系統,會在整個過程中持續調節與校正。我們稍後會看到,它們的角色截然不同卻同樣關鍵。
運動皮質的地圖:潘菲德的小人
二十世紀中葉,神經外科醫師潘菲德(Wilder Penfield)在為癲癇病人開刀時,用微弱電流刺激暴露的運動皮質,記錄病人身體哪個部位產生抽動。他發現 M1 上的神經元呈體感拓樸排列(somatotopic organization):刺激內側引發下肢動作,往外側依序是軀幹、手臂、手、臉。
這張地圖被畫成著名的運動小人(motor homunculus)——一個比例嚴重扭曲的人形。手與臉佔據的皮質面積極大,背部與腿卻很小。這個比例不反映肢體大小,而反映控制精細度:需要精準操作的部位(手指、嘴唇、舌頭)配置了更多神經元。這也解釋了為什麼我們能用手指做出鋼琴演奏這種高解析度動作,卻無法用背部肌肉「寫字」。
不過,現代研究對「一個神經元控制一塊肌肉」的簡化圖像提出了修正。M1 神經元更像是在編碼動作的方向與整體姿態,而非單一肌肉。喬治普洛斯(Apostolos Georgopoulos)在 1980 年代的猴子實驗發現,單一 M1 神經元對特定的伸手方向反應最強,但對鄰近方向也有反應,形成一條調諧曲線(tuning curve)。把同時活動的許多神經元向量加總,就能精準預測手要往哪個方向移動——這就是著名的族群向量(population vector)概念。動作不是由單一神經元決定,而是由神經元族群的集體活動共同編碼。
兩條下行通道:金字塔與金字塔外
從皮質到肌肉,運動指令主要走兩大類路徑。
皮質脊髓束(又稱錐體束,pyramidal tract)是直接通道。大約有一百萬條軸突從運動皮質下行,多數在延腦交叉到對側,因此左腦控制右側身體。這條路徑特別重要的是它對手指獨立、精細動作的控制。當這條束受損(如中風),病人往往失去手部的靈巧度,留下笨拙、不協調的動作。
錐體外系統(extrapyramidal system)則泛指經由腦幹核團(如紅核、前庭核、網狀結構)中繼的下行路徑,主要負責姿勢維持、平衡與大肌群的整體協調。當你站著沒倒、走路時手臂自然擺動,多半是這套系統在背景默默運作。
這裡有一個常見迷思要澄清:很多人以為「運動神經元」就是大腦裡的神經元,但驅動肌肉的下運動神經元(lower motor neuron)其實位於脊髓前角。皮質下來的指令稱上運動神經元(upper motor neuron),它要先「說服」下運動神經元,後者才直接連到肌肉。這個區分在臨床上極重要:上、下運動神經元受損會產生完全不同的症狀(前者肌張力增高、反射亢進;後者肌肉萎縮、反射消失)。
基底核與小腦:兩位風格迥異的協調者
如果說 M1 是「下令的指揮官」,那麼基底核與小腦就是兩位幕後顧問,但他們的工作方式南轅北轍。
基底核:動作的守門人。 基底核是一群皮質下核團(紋狀體、蒼白球、黑質、視丘下核等),它的核心功能可以理解為選擇要執行哪個動作、抑制不該執行的動作。它透過「直接路徑」促進想要的動作,「間接路徑」抑制競爭的動作,形成一種「放行/煞車」的平衡。
帕金森氏症(Parkinson's disease)正是這套系統失衡的經典案例。黑質(substantia nigra)的多巴胺神經元退化,使得「煞車」過強、「放行」不足,病人因此出現動作遲緩(bradykinesia)、僵硬與靜止性顫抖——彷彿想動卻被自己的系統壓制。相對地,亨丁頓舞蹈症(Huntington's disease)則因紋狀體特定神經元退化,導致抑制不足,病人出現無法控制的多餘動作。同一套系統,往兩個方向失衡,產生兩種相反的症狀。
小腦:動作的校準師。 小腦不發動動作,而是比對「實際發生」與「預期發生」之間的誤差,並據此微調。它接收來自運動皮質的「我打算這樣動」副本(efference copy),同時接收來自肌肉、關節的感覺回饋,計算兩者落差後送出修正訊號。
小腦受損的病人會出現運動失調(ataxia):動作的方向大致正確,但時序、力道與終點精度全失準。典型的測試是「指鼻試驗」——要病人用食指碰自己的鼻尖,小腦失調者會在接近目標時出現越來越大的抖動(意向性顫抖),或衝過頭再修正。這顯示小腦的工作是即時的誤差校正與動作平滑化,而非規劃動作本身。
看一個例子:學騎腳踏車時,你的小腦在做什麼
第一次學騎車,你的動作生硬、過度修正——車身一歪,你猛打龍頭,結果矯枉過正往另一邊倒。這時你的運動皮質在「逐一思考」每個動作,小腦的內部模型(internal model)還沒建立。
隨著練習,小腦逐漸學會「給定這樣的傾斜,需要這麼多的修正力道」這個對應關係。它建立起一個前饋模型(forward model):能在感覺回饋還沒到達前,就預測動作的後果並提前修正。這就是為什麼熟練後騎車變得「自動」——你不再需要意識介入,因為小腦已把校正內化成快速、平順的預測性控制。
這個過程也說明了運動學習(motor learning)的本質:不是記住一連串肌肉指令,而是校準大腦對自己身體與外在世界的內部模型。當你拿起一支比平常重的球拍,前幾揮會打偏,因為內部模型還按舊重量預測;幾次之後小腦更新模型,動作又精準了。
反射與脊髓:不必勞煩大腦的捷徑
並非所有動作都要經過大腦。脊髓反射(spinal reflex)是最快的運動迴路。當醫師敲你的膝蓋,肌牽張反射(stretch reflex)讓小腿瞬間踢出——感覺神經偵測到肌肉被拉長,訊號進入脊髓後直接激活運動神經元,不必上達大腦。這條只有一個突觸的迴路(單突觸反射),是已知最快的神經反應之一。
反射的存在揭示了一個重要原則:神經系統把控制分散到最低適當的層級。需要快速、固定回應的事(縮回被燙到的手),交給脊髓;需要彈性判斷的事(要不要接住杯子),才動用皮質。大腦並非事必躬親的微觀管理者,而是把例行工作下放、自己專注於規劃與決策。
動手試試:感受你的本體感覺
閉上眼睛,把雙手舉到身體兩側,再慢慢讓兩手的食指尖在身體前方相碰。多數人能輕鬆做到——即使看不見,你也「知道」手在哪裡。
這種對身體位置的內在感知叫本體感覺(proprioception),來自肌肉、肌腱與關節裡的感受器。運動系統極度依賴它:少了本體感覺回饋,動作會變得笨拙不堪。曾有一位失去本體感覺的著名病例(神經學文獻中的 Ian Waterman),他必須全程用視覺盯著自己的四肢才能移動,一旦關燈就完全無法控制身體。這提醒我們:流暢的動作不只靠「下命令」,更靠持續不斷的「感覺回饋」閉環。
重點回顧
- 運動系統是階層且平行的:高層(前額葉、頂葉)規劃意圖,中層(前運動皮質、輔助運動區)轉成動作序列,低層(初級運動皮質 M1)下達執行指令。
- M1 以族群編碼動作方向:單一神經元有調諧曲線,動作由神經元族群的「族群向量」集體決定,而非一個神經元對一塊肌肉。
- 基底核與小腦角色互補:基底核負責「選擇/抑制」動作(帕金森、亨丁頓是其失衡),小腦負責「誤差校正與平滑化」(受損導致運動失調)。
- 運動學習是更新內部模型:練習的本質是校準大腦對身體與環境的前饋預測模型,而非死記肌肉指令。
- 控制分散到最低適當層級:脊髓反射處理快速固定回應,大腦專注規劃;流暢動作依賴本體感覺的持續回饋閉環。
深入探討(研究所視角)
最適回饋控制理論(optimal feedback control)。 當代運動控制的主流理論框架,由托多羅夫(Emanuel Todorov)與喬丹(Michael Jordan)等人提出,主張運動系統並非追求「精確執行某條預定軌跡」,而是在容許範圍內只修正會影響任務目標的誤差,對不影響目標的變異則放任不管。這解釋了著名的「不受控流形(uncontrolled manifold)」現象——重複同一動作時,與任務無關的維度變異很大,與任務相關的維度卻高度穩定。這個觀點把運動控制重新定義為一個隨機最優化問題,而非開環式的軌跡播放,與工程控制理論深度接軌。
前饋模型與貝氏整合。 小腦的內部模型可用貝氏推論(Bayesian inference)形式化:大腦把前饋預測(先驗)與實際感覺回饋(似然)加權整合,得出對身體狀態的最佳估計(後驗)。當感覺回饋不可靠(如黑暗中),系統更倚重預測;回饋可靠時則更新模型。這套架構不僅解釋了運動控制,也與感覺處理、知覺甚至更廣義的預測編碼(predictive coding)理論相通——大腦或許是一台無時無刻不在預測並修正誤差的機器。
腦機介面與族群解碼。 族群向量的概念已從理論走向應用。透過植入 M1 的電極陣列讀取神經元族群活動,再以解碼演算法(如卡爾曼濾波、遞迴神經網路)即時推估使用者的運動意圖,癱瘓病人已能用「想」的方式操控機械手臂、游標甚至重新書寫文字。這條從喬治普洛斯基礎研究到臨床轉譯的路徑,是基礎神經科學如何長成救命科技的典範。
跨領域連結:與 AI 的雙向啟發。 運動學習與機器學習中的強化學習(reinforcement learning)有深刻對應——基底核的多巴胺訊號被認為編碼「獎賞預測誤差(reward prediction error)」,這正是時序差分學習(temporal difference learning)演算法的核心,舒爾茨(Wolfram Schultz)的猴子實驗為此提供了直接的神經證據。另一方面,小腦的前饋模型啟發了機器人控制中的模型預測控制(model predictive control)。從優生物的角度,肌肉骨骼系統的生物力學限制(如赫胥黎的肌肉滑動細絲理論)決定了神經指令能實現的動作空間;從優心理的角度,動作的自動化與意識控制之間的轉換,正是技能習得與專家表現研究的核心議題。理解運動系統,因此不只是理解「如何動」,更是理解大腦、身體與環境如何在演化與學習中達成精妙協調的縮影。