
同樣一片皮質,憑什麼分得出「視覺區」和「語言區」?發育的真正難題在這裡
從原型圖之爭、人腦的橫向擴張到關鍵期的分子煞車,深入皮質如何把自己切成功能分區,並在睜眼前就完成預訓練。
同樣一片皮質,憑什麼分得出「視覺區」和「語言區」?發育的真正難題在這裡
入門篇帶你走過了大腦發育的主線:神經管捲起、神經元誕生、長途遷徙、軸突尋路、突觸過度生產再被修剪。如果你都記住了,恭喜——但你可能也在心裡留下了一個沒被回答的疑問。
新生的大腦皮質(cerebral cortex)剛長出來時,表面看起來大同小異:六層結構、同樣的神經元類型、相似的局部連線。然而成熟後,枕葉那一小塊變成了處理視覺的初級視覺皮質(V1),顳葉某處變成了理解語言的區域,額葉前端則成為運籌帷幄的前額葉。問題是:一片看似均質的細胞板,到底是「天生就分好區」,還是「被輸入訊號後天教出來」的? 這個被稱為「原型圖 vs 原型皮質(protomap vs protocortex)」的爭論,是發育神經科學最深刻的辯論之一,也是這篇進階文章的起點。
我們將不再重述「大腦會發育」這件事,而是走進三個入門篇刻意略過的難題:皮質如何靠分子座標把自己切成功能分區、人類大腦憑什麼能長得這麼大又這麼皺、以及「關鍵期」這扇窗到底是被什麼分子親手打開又關上的。
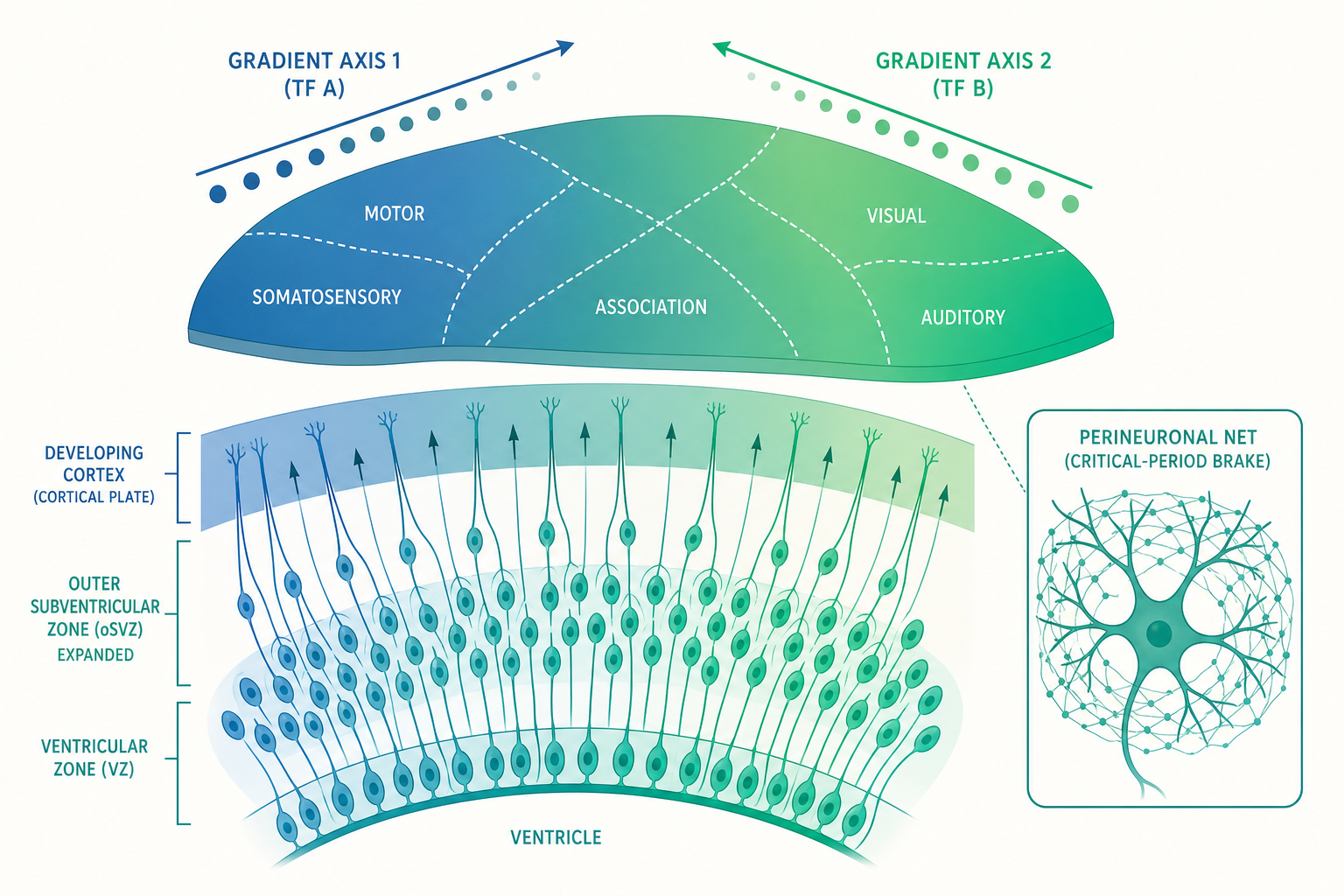
皮質分區的真正機制:原型圖 vs 原型皮質
讓我們先把爭論的兩端講清楚。
「原型皮質假說(protocortex hypothesis)」主張:皮質一開始是一張空白畫布,所有區域的命運取決於它接收到什麼輸入。把眼睛的訊號接到顳葉,顳葉就會學著處理視覺。換句話說,功能分區是經驗與輸入「由外而內」教出來的。
「原型圖假說(protomap hypothesis)」則由 Pasko Rakic 提出,主張:皮質的分區藍圖在神經元還待在腦室區(ventricular zone)的幹細胞階段就已經部分寫定——神經幹細胞本身就帶著「你將來屬於哪一區」的分子身分證,這份「原型圖」隨遷徙被一路帶到皮質表層。
當代的答案是——兩者都對,而且分工精彩。內在的分子梯度先畫出一張「粗略草圖」,再由輸入活動把邊界「精修」清楚。這個整合版的機制核心,是皮質板上同時運作的幾股轉錄因子(transcription factor)梯度。
在發育中的皮質前後(anterior–posterior)與內外(medial–lateral)軸上,幾個關鍵轉錄因子像兩條交叉的座標尺:Pax6 在前外側高、後內側低,Emx2 則剛好相反,後內側高、前外側低,再加上 Coup-TFI(Nr2f1) 與 Sp8 等。每個幹細胞「讀取」自己身上這幾個因子的相對濃度組合,就像讀經緯度一樣,據此決定自己將來屬於運動區、感覺區還是視覺區。實驗證據很漂亮:在小鼠身上人為改變 Emx2 的劑量,整張皮質的功能地圖會等比例地平移與縮放——調高 Emx2,視覺皮質(後內側)變大、運動皮質縮小;調低則相反。這證明分區邊界確實由分子梯度設定,而非單純由輸入決定。
而這些梯度又是誰建立的?答案是皮質邊緣的幾個訊號中心(signaling centers):前端的 FGF8 分泌中心是定義「前方」的關鍵——把額外一團 FGF8 移植到皮質後端,竟能誘導出一個鏡像的、重複的體感區,等於憑空多畫了半張地圖。這呼應了入門篇提到的、沿神經管的 SHH 與 BMP/Wnt 梯度邏輯,只是現在尺度縮小到了單一片皮質內部。
這對「優生物」的學習者是一個重要範式:複雜的空間圖案,不需要一張詳細的「每個細胞的施工藍圖」,只需要少數幾個分子的濃度梯度,加上細胞「讀取局部濃度並做決定」的能力,就能自組織出精密結構。這正是發育生物學家 Lewis Wolpert 著名的「位置資訊(positional information)」與「法國國旗模型」的核心精神——一個你會在「優生物」形態發生(morphogenesis)章節反覆遇到的概念。
人腦憑什麼這麼大?oSVZ 與「橫向擴張」的演化把戲
入門篇說人類大腦約有八百六十億個神經元,並用「放射狀單位假說(radial unit hypothesis)」解釋皮質如何由內而外分層。但這帶出一個演化謎題:人類皮質的神經元數量與表面積遠超老鼠、甚至遠超體型相近的靈長類近親。多出來的神經元,是從哪裡冒出來的?
關鍵不在「分層更多」(哺乳類皮質普遍是六層),而在表面積的橫向擴張。回到放射狀單位假說:每一個腦室區的幹細胞,會生成一「柱」垂直排列的神經元(一個放射狀單位)。柱有多少根,決定皮質鋪開來有多寬;每根柱有多高,才決定厚度。人類之所以擁有巨大的皮質表面積,是因為我們的祖先大幅增加了「柱的數量」——也就是擴張了負責生產這些柱的祖細胞庫。
而這場擴張的主舞台,是一個在人類與其他靈長類特別發達的區域:外側腦室下區(outer subventricular zone, oSVZ)。這裡住著一種關鍵細胞——外側放射狀膠細胞(outer radial glia, oRG 或 bRG)。和傳統貼在腦室壁的放射狀膠細胞不同,oRG 細胞「脫離」了腦室壁,漂浮在更外層,卻保留了強大的自我增殖能力。它們像在工廠外又開了好幾條分廠生產線,使祖細胞庫指數級擴張,因而能供應龐大的神經元數量。在小鼠這層幾乎不存在,在人類卻厚實而活躍——這被認為是靈長類、尤其人類皮質擴張的細胞學基礎。
更引人入勝的是分子層面。近年發現了若干人類特有(human-specific)或人類加速的基因,它們恰好作用在祖細胞增殖這一步:
- ARHGAP11B:一個人類特有、由基因部分複製產生的基因。當研究者把它導入小鼠胚胎,竟能促使小鼠皮質祖細胞擴增,甚至誘導出本來不該有的腦回皺褶——一個基因,撥動了「腦變大變皺」的開關。
- NOTCH2NL 家族:人類特有的 Notch 訊號調控基因,能延長神經祖細胞停留在「持續分裂」狀態的時間,從而產生更多神經元。
這些發現把「人類為何擁有獨特認知能力」的問題,第一次部分地落實到具體的、可實驗操弄的分子上。當然,我們要謹慎:擁有更多神經元不等於更聰明(鯨豚與大象的某些腦區神經元也很多),認知能力還牽涉連結模式、髓鞘化時程與發育延長(人類童年特別長)。但 oSVZ 與這些基因,至少讓「人類大腦的演化」從一個哲學命題,變成了實驗台上可解的生物學問題。
看一個例子:腦類器官如何「在培養皿裡」重演這段歷史
要研究人類專屬的發育機制,有個棘手的倫理與技術障礙:你不能拿人類胎兒大腦做實驗,而小鼠又恰好缺了 oSVZ 這層最關鍵的構造。
過去十年,腦類器官(brain organoid)技術提供了一條出路。研究者把人類的誘導性多能幹細胞(induced pluripotent stem cells, iPSC)放進特定的訊號環境中培養,這些細胞會自發地(self-organize)形成帶有腦室區、分層神經元、甚至可辨識出 oRG 細胞與 oSVZ 樣結構的三維組織——一顆毫米級、會放電的「迷你大腦」。
這帶來幾個強大的應用。第一,比較演化:用人類、黑猩猩、獼猴的細胞各做一批類器官並排比較,研究者能精準找出「人類版本」在祖細胞分裂、神經元產量上的差異,再回去定位是哪些基因(如前述 ARHGAP11B)造成的。第二,疾病建模:帶有特定突變(如導致小頭畸形 microcephaly 的基因)的病人 iPSC 做出的類器官,會重現異常縮小的尺寸,讓研究者在培養皿裡「看見」疾病如何在發育早期成形。第三,致畸原研究:2015–2016 年茲卡病毒(Zika virus)疫情期間,正是腦類器官實驗迅速證實了病毒會優先感染並殺死神經祖細胞,從機制上解釋了它為何導致新生兒小頭畸形。
當然,類器官遠非真正的大腦——它缺乏血管、缺乏感覺輸入、也沒有完整的區域組織與成熟迴路,把它擬人化是危險的。但作為「在受控條件下重演人類早期皮質發育」的工具,它已經成為這個領域最重要的方法學突破之一,也是「優生物」與幹細胞工程交會的前沿。
關鍵期不是時鐘自己走完的:抑制性神經元與分子煞車
入門篇用 Hubel 與 Wiesel 的視覺剝奪實驗,介紹了「關鍵期(critical period)」——大腦某些迴路只在特定時間窗對經驗高度敏感,錯過就難以重塑。入門篇的「深入探討」也預告了:關鍵期的開與關是被主動控制的。現在,我們把這個機制徹底打開。
長久以來,人們直覺地以為關鍵期像一個內建的鬧鐘:時間到了自動開、時間到了自動關,由某種發育時鐘被動倒數。但 Takao Hensch 等人的研究徹底改寫了這個圖像:關鍵期的開啟,由一類特定神經元的成熟所「點火」;它的關閉,則由一套分子「煞車」主動鎖上。 換言之,關鍵期不是被動的時間流逝,而是被主動開關的生理狀態。
點火的關鍵,是一類抑制性中間神經元——含小白蛋白的 GABA 能中間神經元(parvalbumin-positive, PV interneurons)。它們的成熟,調整了皮質中「興奮與抑制的平衡(excitation/inhibition balance, E/I balance)」。在 PV 神經元尚未成熟前,皮質迴路的抑制太弱、訊號太雜,無法形成清晰可被經驗雕刻的活動模式;當 PV 神經元成熟、抑制達到某個臨界水準,迴路才進入「可被經驗精準塑形」的狀態——關鍵期於是開啟。最直接的證據是:用藥物(如苯二氮平類 benzodiazepine)人為提早增強 GABA 抑制,可以提早啟動視覺皮質的關鍵期;而在缺乏正常 GABA 合成的小鼠身上,關鍵期則遲遲不開,直到補上抑制才啟動。
那麼關閉呢?關鍵期之所以會結束,是因為一套「結構性煞車」逐漸到位,把可塑性鎖死。其中最具代表性的是神經周圍網(perineuronal nets, PNNs)——一層由聚醣(chondroitin sulfate proteoglycans)構成、像漁網一樣包覆在成熟 PV 神經元周圍的細胞外基質。PNN 一旦形成,等於把這些神經元的連結「上了水泥」,大幅限制了突觸的增減。漂亮的反證是:用一種酵素(chondroitinase ABC)在成年動物身上消化掉 PNN,竟能重新開啟一段本該早已關閉的可塑性窗口——這直接呼應了入門篇預告的「重新開啟關鍵期」的可能性。此外,髓鞘相關的抑制訊號(如 Nogo、MAG)與 Lynx1 等分子,也都是在發育後期築起、限制可塑性的煞車之一。
把這套機制連起來看,會得到一個深刻的觀念:「可塑性」與「穩定性」是一對需要被主動權衡的對立量。 幼年大腦選擇了高可塑性以便快速學習與適應,代價是迴路不穩、容易被干擾;成熟大腦則選擇用 PNN 等煞車鎖住辛苦學成的迴路,換取穩定與效率,代價是學新東西變慢。這不是缺陷,而是一場有意識的工程取捨——而且這把鎖,原則上是可以被分子鑰匙重新打開的。
在經驗到來之前:大腦如何「自己練習」連線
還有一個入門篇沒談、卻顛覆直覺的面向:有些迴路的精修,在感覺經驗根本還沒開始之前就完成了。 我們很容易以為「經驗雕刻迴路」意味著要等到睜開眼、聽到聲音之後才發生。但大腦遠比這聰明——它會在出生前「自己製造假輸入」來預先調校。
最經典的例子是視覺系統的自發視網膜波(spontaneous retinal waves)。在胎兒尚未睜眼、視網膜還無法感光時,視網膜上的神經節細胞會自發地、像漣漪一樣一波波橫掃地集體放電。這些「波」不是雜訊,而是高度結構化的活動:相鄰的細胞會同步、距離遠的則不同步。
回想入門篇提到、可塑性進階篇深入過的赫布原則(「一起激發的連在一起」)。視網膜波恰好為這條原則提供了「練習用的素材」:因為相鄰視網膜細胞傾向同步放電,它們投射到視丘與視覺皮質的軸突也會同步活動,於是「來自視網膜相鄰位置」的連結被一起強化、保留;而隨機不同步的連結被削弱。結果是:在動物還沒看過任何真實影像之前,視覺通路就已經建立好了精準的『地形對應(retinotopic map)』——讓視野相鄰的點,對應到皮質相鄰的位置。 同樣的自發活動機制,也參與了「眼優勢柱(ocular dominance columns)」這種把左右眼輸入分流成交替條紋的精巧結構的初步建立。
這帶出一個對「優心理」與「AI」都極具啟發的觀念:先天與後天的界線,比我們想像的更模糊。 視覺迴路的初步骨架,既不是純粹寫死在基因裡(它需要神經活動來雕刻),也不是來自真實的外界經驗(眼睛還沒睜開)——它來自大腦內生的、自我產生的活動模式。大腦不是被動等待世界來教它,而是先用「內部模擬」把自己預訓練(pre-train)好,等真實輸入一到便能立刻上手。這幾乎就是當代 AI「自監督預訓練(self-supervised pre-training)」的生物版本:在拿到標註資料之前,先用大量自我生成的結構把網路調到一個好的起點。
重點回顧
- 皮質分區是「原型圖」與「輸入」的合作:Pax6、Emx2 等轉錄因子梯度(由 FGF8 等訊號中心建立)先畫出粗略功能地圖,再由輸入活動精修邊界——改變一個分子劑量就能平移整張腦地圖。
- 人腦的大,來自橫向擴張而非分更多層:外側腦室下區(oSVZ)的外側放射狀膠細胞(oRG)大幅擴增祖細胞庫;ARHGAP11B、NOTCH2NL 等人類特有基因把這件事落實到可實驗的分子層次。
- 腦類器官讓人類專屬發育可被實驗:用 iPSC 自組織出帶 oSVZ 樣結構的迷你腦,用於演化比較、疾病建模與致畸原(如茲卡病毒)研究,但它不是真正的大腦。
- 關鍵期是被主動開關的,不是時鐘走完:PV 抑制性神經元成熟(調整 E/I 平衡)點火開啟,神經周圍網(PNN)等分子煞車鎖上關閉;消化 PNN 可在成年重啟可塑性。
- 經驗到來前大腦已自我預訓練:自發視網膜波等內生活動,在睜眼前就靠赫布機制建好了地形對應與眼優勢柱的雛形——這是「自監督預訓練」的生物原型。
深入探討(研究所視角)
第一,從「原型圖」到精準醫療:發育時序的因果重建。 過去我們只能在固定的死後組織上做「快照」,難以追蹤單一細胞如何隨時間改變身分。如今單細胞 RNA 定序(single-cell RNA-seq)搭配譜系追蹤(lineage tracing)與空間轉錄體學(spatial transcriptomics),讓研究者能在發育的連續時間點上,同時掌握「每個細胞此刻表達哪些基因」與「它身處皮質哪個位置」。這正在把 Pax6/Emx2 那張靜態梯度圖,升級成一部動態的「細胞命運決策影片」。對精神疾病研究而言意義重大:若思覺失調症或自閉症源於發育早期的細微偏差(入門篇深入探討已提及此「神經發育假說」),那麼唯有具備時間解析度的工具,才能定位偏差究竟發生在哪個時間窗、哪群細胞。
第二,類器官的倫理前沿與 Ethicomics。 腦類器官的能力正在逼近一些令人不安的問題:當類器官變得更大、開始出現自發的、類似早產兒腦波的協調電活動時,它是否可能具有任何形式的「感受」?目前主流共識是現有類器官遠不具備意識所需的複雜度與整合度,但學界已主動展開「類器官倫理(organoid ethics)」的討論,預先設定研究紅線。這正是 Educational Omics 框架中 Ethicomics 維度的真實案例:科學能力跑在前面時,倫理規範必須主動、而非被動地跟上。對在 Uedu 跨領域學習的同學,這是一個極好的反思題材——技術的「能做」與「該做」之間,永遠需要人為的審議。
第三,可塑性煞車的臨床轉譯:把成年大腦調回「幼年模式」。 關鍵期煞車的分子機制(PNN、E/I 平衡、Lynx1 等)一旦被解析,自然引出一個誘人的治療方向:在中風復健、弱視(amblyopia)治療、甚至成人技能學習中,能否暫時、局部地「鬆開煞車」,重啟一段可塑性窗口?已有動物實驗顯示消化 PNN、或調控 E/I 平衡能恢復成年可塑性,部分藥物(如某些抗憂鬱劑)也被發現能短暫重啟皮質可塑性。但這把雙面刃必須謹慎:可塑性是一種權衡,過度鬆開煞車可能讓辛苦穩定的迴路重新變得脆弱、易受干擾。「該不該、以及如何精準地重開一扇關鍵期窗口」,是發育神經科學最具應用潛力、也最需要節制的前沿。
最後,發育作為 AI 的設計藍圖。 本文兩處(自發視網膜波的「自監督預訓練」、關鍵期的「可塑性—穩定性權衡」)都指向同一個跨領域洞見:大腦的發育策略,正在反過來啟發人工智慧的架構設計。AI 工程師早已借用「課程學習(curriculum learning)」——讓模型先學簡單再學難,呼應發育的階段性;也借用「可塑性—穩定性兩難(plasticity–stability dilemma)」這個直接源自神經科學的詞彙,來描述持續學習系統「既要學新、又不能忘舊」的困境。更前沿的方向是探索「生長式(developmental / growing)神經網路」:不再從一個固定大小的網路開始訓練,而是讓網路像皮質一樣,隨任務需求動態增生與修剪結構。對於想橫跨「優神經科學」與「AI」的你,神經發育提供的不只是比喻,而是一整套經過數億年演化驗證、關於「如何讓一個系統從簡單規則自我組織出複雜智能」的工程原理。讀懂大腦怎麼長出來,或許正是讀懂下一代人工智慧該怎麼「長」的起點。