
倫敦計程車司機的海馬迴,為什麼比你大?
從赫布定律、LTP/LTD 到關鍵期與損傷重組,理解大腦如何隨經驗物理性地改寫自身結構。
倫敦計程車司機的海馬迴,為什麼比你大?
要拿到倫敦的計程車駕照,你得先通過一場被稱為「知識大全(The Knowledge)」的恐怖考試:把市中心約兩萬五千條街道、上千個地標的相對位置全部記進腦袋,能在任意兩點之間即時規劃路線。準備期平均三到四年。神經科學家 Eleanor Maguire 在 2000 年掃描了這群司機的大腦,發現了一件驚人的事——他們大腦中負責空間記憶的「海馬迴後部(posterior hippocampus)」,灰質體積明顯比一般人大;而且年資越久、跑得越熟,這塊區域就越大。後續追蹤更顯示,那些通過考試的見習司機,海馬迴是在訓練「之後」才長大的,沒通過的人則沒有變化。
這意味著什麼?意味著你此刻所經歷的每一次學習、每一段記憶、每一個反覆練習的動作,都在悄悄地、物理性地改寫你大腦的結構。大腦不是一台出廠後規格固定的硬體,而是一塊終身可被經驗雕塑的黏土。這種能力,就是我們今天的主角——神經可塑性(neuroplasticity)。
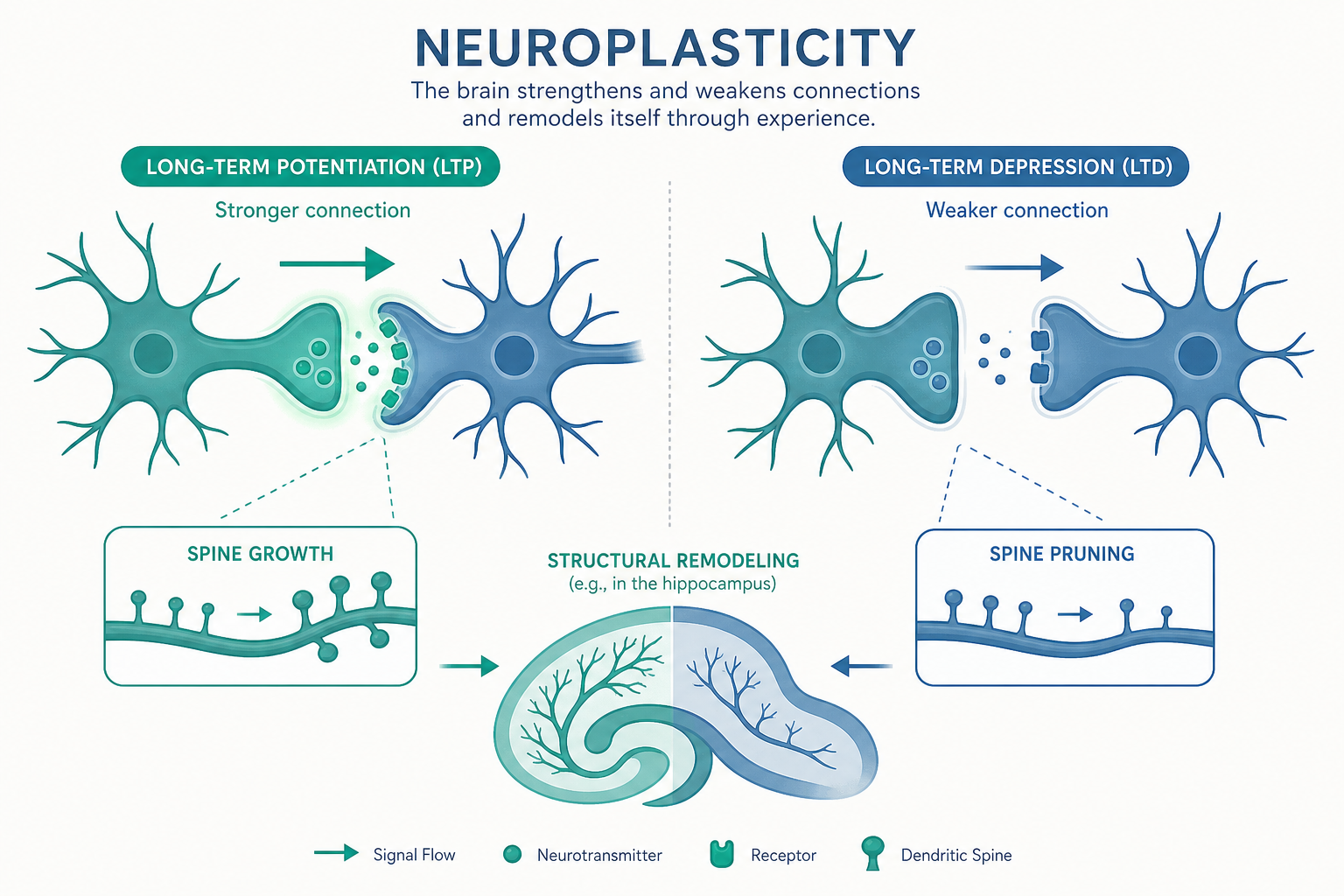
可塑性不是比喻,而是分子層級的真實事件
「大腦會改變」聽起來像勵志口號,但神經可塑性是一個有明確物理基礎的科學概念。它指的是神經系統在結構與功能上,因應經驗、學習、發育或損傷而發生改變的能力。這種改變可以小到一個突觸(synapse)的強度微調,也可以大到整片皮質區域的功能重新分配。
要理解它,我們得先回到神經元(neuron)的連接點。兩個神經元之間並不直接相連,而是隔著一道極窄的縫隙——突觸。上游神經元(突觸前)釋放神經傳導物質(neurotransmitter),下游神經元(突觸後)的受體接收訊號。可塑性的核心,就發生在這個介面上。
神經科學家把可塑性大致分成兩個層次:
- 功能性可塑性(functional plasticity):突觸傳遞效率的改變。同樣的訊號傳過來,這個突觸現在讓下游反應得更強或更弱。這發生得很快,幾分鐘到幾小時。
- 結構性可塑性(structural plasticity):實際的物理結構改變——長出新的樹突棘(dendritic spine)、新的突觸、軸突分支重新走向,甚至(在特定腦區)產生全新的神經元。這發生得較慢,數天到數月。
倫敦司機的海馬迴變大,屬於結構性可塑性的宏觀表現;而你背一個英文單字時當下發生的,多半是功能性可塑性。
赫布定律:一起激發的神經元,會連在一起
要講可塑性的機制,繞不開一句神經科學界最著名的格言。1949 年,心理學家 Donald Hebb 提出一個假設,後來被濃縮成一句口訣:
「Cells that fire together, wire together.」(一起激發的細胞,會連結在一起。)
赫布的洞見是:如果神經元 A 持續、反覆地參與激發神經元 B,那麼 A 與 B 之間的連結就會被強化。這就是學習的細胞層級基礎。當你反覆把「apple」這個字和「蘋果」這個概念一起激活,承載這兩者的神經元群之間的突觸就被一次次加強,最後變得一觸即發。
但赫布定律有個常被忽略的下半句:沒有一起激發的,連結會被削弱。可塑性是雙向的——強化(potentiation)與削弱(depression)並存。這非常重要,因為如果突觸只能變強不能變弱,大腦很快就會飽和、失控。學習不只是「記住」,也包括「修剪掉」不相關的連結。
LTP 與 LTD:可塑性的分子引擎
赫布的假設在 1973 年得到實驗證據。Bliss 與 Lømo 在兔子海馬迴中發現:用一串高頻電刺激轟擊一條神經通路後,這條通路的突觸傳遞效率會持續增強好幾個小時甚至更久。這個現象被命名為長期增益(long-term potentiation, LTP),至今仍是學習與記憶的主流細胞模型。
LTP 的分子機制裡有個明星角色:NMDA 受體(NMDA receptor)。它是一個「巧合偵測器(coincidence detector)」——只有在兩個條件同時成立時才會開啟:
- 突觸前神經元釋放了麩胺酸(glutamate);
- 突觸後神經元本身已經被去極化(depolarized,也就是已經處於興奮狀態)。
當兩者同時發生,NMDA 受體的通道才打開,讓鈣離子(Ca²⁺)大量湧入突觸後細胞。鈣離子是關鍵的細胞內訊使,它啟動一連串級聯反應,最終讓更多 AMPA 受體被插入突觸後膜——突觸因此變得對訊號更敏感。這完美對應了赫布定律:「突觸前活動」加上「突觸後活動」同時發生,連結才被強化。
它的反面是長期減弱(long-term depression, LTD):當突觸前後的活動只有微弱、不同步的關聯時,鈣離子流入較少、較緩,反而觸發另一套訊號路徑,把 AMPA 受體從突觸移除,使連結變弱。LTP 與 LTD 一進一退,讓大腦能精細地調校每一條連結的權重。
看一個例子:學騎腳踏車與「程序記憶」的固化
想想你學騎腳踏車的過程。一開始你搖搖晃晃,得用力思考每個動作——重心、踏板、把手。這時參與運作的腦區很廣,前額葉(prefrontal cortex)忙著用意識控制每個細節,效率低又費力。
隨著一次次練習,掌管動作協調的小腦(cerebellum)與基底核(basal ganglia)中的相關突觸被反覆活化,依循 LTP 機制逐漸強化。同時,那些「多餘」的、導致你跌倒的錯誤連結,透過 LTD 被削弱、修剪掉。神經迴路從「廣而散」收斂成「窄而專」。
幾週後,你不再需要思考就能騎。控制權從耗能的前額葉,轉移到了高度自動化的皮質下迴路。這就是為什麼騎車一旦學會幾乎不會忘——這份「程序記憶(procedural memory)」已經被結構性可塑性穩固地寫進了你的運動迴路。這也解釋了一個現象:你可以十年沒碰腳踏車,跨上去還是會騎。那條被雕刻好的神經路徑,依然在。
關鍵期:為什麼有些窗口會關上
可塑性並非終身均勻。大腦發育有所謂關鍵期(critical period)——某些功能在特定時間窗內可塑性極高,過了窗口就大幅下降。
最經典的證據來自 Hubel 與 Wiesel 的視覺系統研究(他們因此獲得 1981 年諾貝爾生理醫學獎)。他們在幼貓發育早期暫時遮住一隻眼睛,發現視覺皮質中原本應該接收那隻眼睛訊號的區域,會被另一隻眼睛「搶走」。更關鍵的是:如果遮蔽發生在關鍵期內,影響是永久的;但若在成年後才遮同樣久的時間,視覺皮質幾乎不受影響。這對應到人類臨床上的「弱視(amblyopia)」——必須及早矯正,錯過窗口就難以恢復。
語言學習也有類似的窗口。母語的語音辨識能力在嬰兒期極具彈性,這也是為什麼幼年接觸第二語言通常能達到母語般的口音,而成年後學外語往往帶腔。但這裡要破除一個迷思:關鍵期關上不代表可塑性歸零。成年大腦依然高度可塑——只是需要更多的努力、重複與專注,而且某些底層迴路(如基礎語音區辨)的彈性確實大幅降低。倫敦計程車司機都是成年人,他們的海馬迴照樣長大了。
損傷後的重組:大腦的自我修復
可塑性最戲劇性的展現,是在腦損傷之後。中風患者損傷了控制右手的左腦運動區,初期右手癱瘓;但透過密集復健,許多人能逐步恢復部分功能。這不是受損的神經元復活了,而是周邊未受損的腦區接管了原本的功能——鄰近皮質、甚至對側半球重新組織迴路,把失去的功能「分包」出去。
這背後的臨床原理催生了一種療法:限制誘發運動療法(constraint-induced movement therapy, CIMT)。做法是把患者「好的那隻手」綁起來,強迫他大量使用患側的手。違反直覺,卻有效——因為這逼迫受損半球周邊區域被密集活化,誘發可塑性重組。這正是「用進廢退」的可塑性原則在臨床上的落實:被使用的迴路被強化,被閒置的迴路被佔用。
重點回顧
- 神經可塑性是物理真實:學習與經驗會實際改變突觸強度與大腦結構,倫敦計程車司機變大的海馬迴是經典證據,而非比喻。
- 赫布定律是核心法則:「一起激發的神經元會連在一起」——但別忘了下半句,沒一起激發的連結會被削弱,可塑性是雙向的。
- LTP 與 LTD 是分子引擎:NMDA 受體作為「巧合偵測器」,透過鈣離子訊號決定突觸是被強化(LTP)還是削弱(LTD)。
- 關鍵期讓某些窗口會關上:視覺、語言等功能在發育早期可塑性最高,但成年大腦依然高度可塑,只是需要更多重複與專注。
- 損傷後可重組:未受損腦區能接管失去的功能,CIMT 等復健療法正是利用「用進廢退」的可塑性原則。
深入探討(研究所視角)
穩定與彈性的兩難:恆定可塑性(homeostatic plasticity)
純粹的赫布式可塑性有個理論上的致命缺陷:它是正回饋。強的連結被用得更多、變得更強、又被用得更多——這會導致活動失控地爆衝或徹底沉寂。大腦如何避免這場災難?答案是恆定可塑性,特別是 Gina Turrigiano 提出的「突觸縮放(synaptic scaling)」機制。當神經元整體活動長期偏高,它會「按比例」調降所有突觸的強度;偏低則整體上調。這套全域性的慢速調節,與快速、突觸專一的赫布式可塑性協同運作,讓大腦既能學習(彈性)又不失穩定。理解這組張力,是當代可塑性研究的核心命題之一。
不只是神經元:膠細胞與神經調節物質
傳統可塑性研究聚焦於神經元,但近年焦點正在擴張。星形膠細胞(astrocyte)並非被動的支持細胞,而會主動參與突觸的形成、修剪與傳遞調節,催生了「三方突觸(tripartite synapse)」的概念。此外,多巴胺(dopamine)、乙醯膽鹼(acetylcholine)等神經調節物質扮演「可塑性的開關」——它們標記「此刻值得學習」的時刻(例如獎賞、新奇、專注),讓 LTP 在這些時刻更容易發生。這正是為什麼「動機」與「注意力」在學習中如此關鍵:它們不只是心理狀態,而是有分子層級的可塑性後果。這條線索直接連結到優心理的動機理論與優生物的神經內分泌調節。
成年神經新生的爭議
成年人腦中是否會長出全新神經元?這是一個尚未完全定論的前沿問題。海馬迴齒狀回(dentate gyrus)的成年神經新生(adult neurogenesis)在囓齒類動物中證據確鑿,且與學習、情緒調節相關;但在人類身上,不同研究團隊用不同方法得出相互矛盾的結論——有的宣稱終身持續,有的宣稱青春期後幾乎停止。這提醒研究者:可塑性的種類與程度在不同物種、不同腦區差異極大,不能簡單外推。對學習者而言,重要的是不要把「成年神經新生」當成已成定論的勵志素材。
跨領域連結:人工智慧的可塑性難題
赫布定律不只啟發了神經科學,也是人工神經網路(artificial neural network)的思想源頭——「調整連結權重以反映共現關係」正是機器學習的核心。但生物與人工系統有個耐人尋味的對比:災難性遺忘(catastrophic forgetting)。當人工神經網路學習新任務時,往往會抹除舊任務的記憶,因為新的權重更新蓋掉了舊的。而人腦卻能持續學習一輩子而不崩潰——這正得益於前述的恆定可塑性、神經調節的選擇性閘控、以及睡眠期間的記憶固化(memory consolidation)等多重機制協同保護舊記憶。如何讓 AI 擁有大腦這種「持續學習(continual learning)」能力,是當前機器學習的活躍前沿。這也是 Uedu 多模態學習分析所關注的:當我們在 AI 與 優神經科學 之間架橋時,生物可塑性既是靈感的來源,也是衡量人工系統的一面鏡子。
對學習實踐的啟示
把這些機制收束回學習現場,幾個原則便顯而易見:可塑性需要重複(單次活化不足以穩固 LTP)、需要間隔(分散練習讓記憶固化得以發生,呼應「間隔效應」spacing effect)、需要睡眠(深層睡眠是突觸重整與記憶固化的關鍵窗口)、也需要主動投入(神經調節物質只在專注與動機在場時開啟可塑性閘門)。你的大腦終身可被雕塑——而你,正握著那把刻刀。