
從一顆細胞到八百六十億個神經元:大腦是怎麼「長」出來的?
從神經管到成熟前額葉,拆解大腦發育的過度生產、精準修剪與經驗雕刻,並理解青少年行為與終身可塑性。
從一顆細胞到八百六十億個神經元:大腦是怎麼「長」出來的?
懷孕第三週,一位準媽媽可能還沒意識到自己懷孕,胚胎背側卻已經悄悄發生了一件大事:一片扁平的細胞板捲起來,封閉成一根中空的管子。這根不起眼的「神經管」(neural tube),就是未來腦與脊髓的全部起點。短短九個月內,它將擴張、摺疊、分化,最終長成一個擁有約八百六十億個神經元(neuron)、數以兆計突觸(synapse)的器官。而這個器官還要再花將近二十五年,才真正成熟。
我們常把「大腦發育」想像成一條從小到大、線性放大的過程,像吹氣球一樣愈吹愈大。但真實的神經發育(neurodevelopment)遠比這精彩,也更違反直覺:它包含了爆炸性的「過度生產」與大規模的「刪減」,包含細胞長途遷徙、軸突精準尋路,也包含環境經驗親手雕刻迴路。理解這段歷程,不只是生物學的好奇,更直接關係到我們如何理解學習、語言、青少年行為,乃至於發展障礙的成因。
第一階段:神經管與大腦的「分區藍圖」
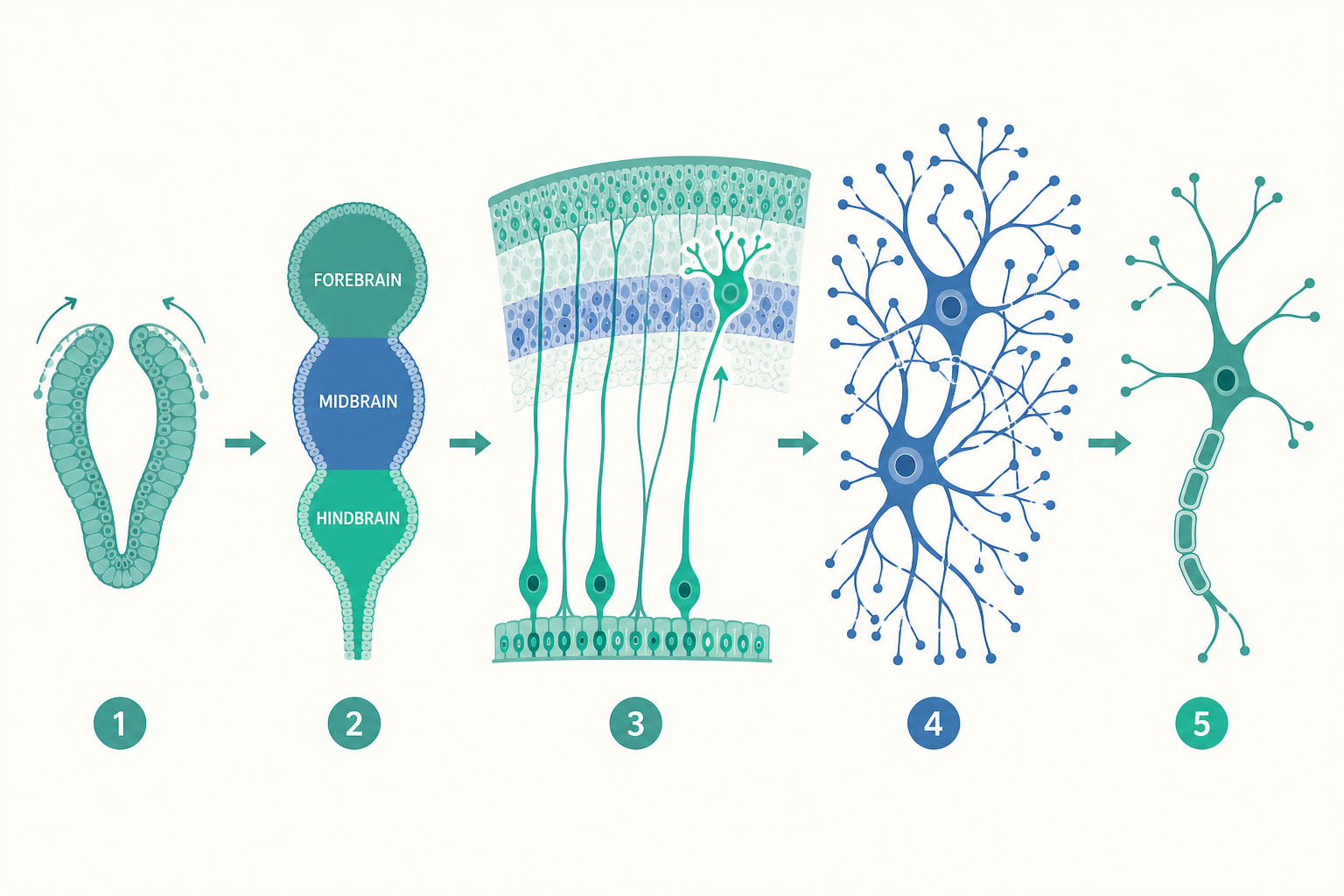
故事從「神經誘導」(neural induction)開始。胚胎早期的外胚層(ectoderm)原本可以變成皮膚,但底下的脊索(notochord)會分泌訊號分子,告訴上方一片細胞:「你們不要當皮膚,去當神經組織。」這個過程的關鍵,是一種叫做 BMP 的訊號被抑制——著名的 Spemann 與 Mangold 在 1924 年的胚胎移植實驗(即「組織者」organizer 的發現)正是這套邏輯的經典證據,後來也讓相關研究獲得諾貝爾獎肯定。
被誘導出的神經板(neural plate)隨後捲合成神經管。神經管前端膨大,依序形成三個原始腦泡:前腦(prosencephalon)、中腦(mesencephalon)、後腦(rhombencephalon),再進一步細分為五個區域,分別對應未來的大腦半球、間腦、中腦、橋腦小腦與延髓。這裡有一個重要觀念:大腦的「分區」不是後來才劃分的,而是在最早期就由分子梯度預先寫好藍圖。沿著神經管的前後軸,Hox 基因家族像門牌號碼一樣標定位置;沿著背腹軸,背側的 BMP/Wnt 與腹側的 Sonic hedgehog(SHH)形成濃度相反的兩道梯度,細胞「讀取」自己所在的濃度,決定要分化成感覺神經元還是運動神經元。
如果這一步出錯,後果嚴重。當神經管前後端未能正常閉合,就會導致無腦畸形(anencephaly)或脊柱裂(spina bifida)。這也是為什麼孕前與孕早期補充葉酸(folic acid)被列為公共衛生重點——它能顯著降低神經管缺陷的風險。發育的「藍圖」雖然由基因主導,卻早早就和營養等環境因素糾纏在一起。
第二階段:神經發生與細胞遷徙——一場有秩序的大遷移
藍圖畫好後,接著要「蓋房子」。在神經管內壁的腦室區(ventricular zone),神經幹細胞(neural stem cells)瘋狂分裂。在人類胎兒發育的高峰期,大腦平均每分鐘要新增數十萬個神經元——這個數字常被引用為「每分鐘約二十五萬個」,用以強調神經發生(neurogenesis)的驚人速度。
但神經元誕生的地方,往往不是它最終工作的地方。以大腦皮質(cerebral cortex)為例,新生的神經元必須從深處的腦室區,沿著一種特化的「放射狀膠細胞」(radial glia)所形成的纖維,像攀岩者抓著繩索一樣,往外遷徙到皮質表層。更精妙的是,皮質是「由內而外」一層層搭建的:最早出生的神經元留在最深層,後出生的神經元要「越過」先到者,停在更外側。這個由 Rakic 提出並驗證的「放射狀單位假說」(radial unit hypothesis),解釋了人類六層皮質如何依序堆疊而成。
當遷徙的導航出錯,皮質的分層就會錯亂,臨床上可見如「無腦回畸形」(lissencephaly,字面意思是「平滑的腦」)——大腦表面失去正常的溝回。許多這類疾病可追溯到特定基因(如 LIS1、DCX)的突變,這些基因正是負責細胞遷徙的分子機器。這提醒我們:大腦複雜的外形,背後是一連串可被單一基因打斷的精密程序。
第三階段:軸突尋路與突觸形成——連線的藝術
神經元就定位後,真正困難的問題才浮現:一個位於運動皮質的神經元,如何把它的軸突(axon)精準延伸到數十公分外的脊髓,連上正確的目標?這在尺度上,相當於一個人從房間出發,要在全黑的城市裡找到指定的某一扇門。
答案是軸突末端有一個像探測器的構造——「生長錐」(growth cone)。它伸出許多細小的觸手,不斷感測環境中的化學訊號。Sperry 在一九四〇至六〇年代以青蛙視神經做的經典實驗提出「化學親和假說」(chemoaffinity hypothesis):軸突與目標細胞帶有彼此匹配的分子「標籤」,靠吸引與排斥訊號(如 Netrin、Slit、Ephrin、Semaphorin 等家族)一段段導航到正確終點。這是神經科學最優雅的概念之一,也呼應了「優生物」中分子辨識與專一性結合的核心邏輯。
軸突抵達目標後,與下一個神經元之間形成突觸。值得注意的是,這個階段大腦採取了一種看似「浪費」的策略:它先製造遠超所需的突觸。人類嬰兒在出生後頭兩年,突觸密度急速攀升,在某些皮質區域,幼兒的突觸數量甚至超過成人。為什麼要造這麼多?答案藏在下一個階段。
第四階段:突觸修剪與髓鞘化——少即是多
如果說前面是「加法」,那麼接下來就是「減法」。出生後,大腦進入一段漫長的「突觸修剪」(synaptic pruning)時期:被頻繁使用、有實際功能的連結會被保留並強化,而閒置、冗餘的連結則被移除。這個過程一路持續到青春期之後。
修剪不是隨機的,它遵循一條深刻的原則——「用進廢退」,在神經科學中常被表述為赫布理論(Hebbian theory):「一起激發的神經元會連在一起」(neurons that fire together, wire together)。Hubel 與 Wiesel 在一九六〇年代以貓的視覺系統做的實驗是最有力的證據:他們在幼貓發育的關鍵時期遮蔽一隻眼睛,結果該眼對應的皮質連結大幅萎縮,即使日後恢復視力也無法挽回。這項研究讓他們獲得一九八一年諾貝爾生理醫學獎,也確立了「關鍵期」(critical period)的概念——大腦有些迴路只在特定的時間窗內對經驗高度敏感,錯過了就難以重塑。
與修剪同時進行的,是「髓鞘化」(myelination)。寡突膠細胞(oligodendrocyte)像電工纏絕緣膠帶一樣,在軸突外包裹一層層富含脂質的髓鞘,讓電訊號傳導速度提升數十倍。髓鞘化從胎兒晚期開始,但前額葉皮質(prefrontal cortex)的髓鞘化最晚完成,要到二十多歲。這個生物事實,正是理解青少年的鑰匙。
看一個例子
讓我們把鏡頭對準一位十六歲的高中生。她在某些時刻展現出驚人的洞察力與創造力,但在另一些時刻又做出衝動、明知有風險仍冒險的決定。家長與老師常感困惑:「她明明很聰明,為什麼還會這樣?」
神經發育給了我們一個不帶評價、卻極具說服力的解釋。青春期的大腦並非「未發育完全的成人腦」,而是一個正處於特殊失衡狀態的系統。主管情緒與獎賞的邊緣系統(limbic system,特別是杏仁核 amygdala 與依核 nucleus accumbens)在青春期受荷爾蒙驅動,發育得又早又活躍,對刺激、新奇與同儕認可的反應格外強烈。然而負責踩煞車的前額葉皮質——掌管衝動控制、後果評估、長期規劃——卻是全腦最晚成熟的區域,它的髓鞘化與突觸修剪要到二十五歲左右才大致完成。
Steinberg、Casey 等研究者提出的「雙系統模型」(dual systems model)正是描述這種「油門早熟、煞車晚熟」的暫時性落差。這不是缺陷,而是演化上可能有意義的設計:青春期的探索傾向有助於個體離開原生家庭、學習新技能、建立社會連結。理解這一點,能讓教育現場從「為什麼這個學生這麼不理性」轉向「我如何設計環境,在煞車尚未裝好前提供合適的支持」。這也與「優心理」中對自我調節(self-regulation)發展的討論直接呼應——調節能力不只是意志問題,更有其神經成熟的時間表。
重點回顧
- 大腦發育不是線性放大,而是「過度生產 + 精準刪減」:神經元與突觸先大量製造,再透過修剪保留有用連結,體現「少即是多」的設計哲學。
- 分區藍圖在最早期就由分子梯度寫定:Hox 基因、SHH、BMP/Wnt 等訊號沿前後軸與背腹軸形成梯度,細胞依所在濃度決定命運。
- 位置決定功能,遷徙不能出錯:皮質「由內而外」分層搭建,遷徙障礙會導致無腦回畸形等結構異常。
- 經驗親手雕刻迴路:Hubel 與 Wiesel 的視覺剝奪實驗證實了「關鍵期」與「用進廢退」(Hebbian)原則,環境經驗在特定時間窗內具有決定性影響。
- 前額葉最晚成熟,解釋青少年行為:油門(邊緣系統)早熟、煞車(前額葉)晚熟的暫時失衡,是發育常態而非缺陷。
深入探討(研究所視角)
進入研究層次,神經發育有幾個值得深思的開放問題。
第一,先天與後天的對立其實是個假問題。 古典爭論常問「智力是基因決定還是環境決定」,但分子發育生物學已清楚顯示,基因從不在真空中運作。SHH 梯度需要細胞「讀取」並轉譯成基因表現,而表現又受到表觀遺傳(epigenetic)調控——DNA 甲基化、組蛋白修飾等機制讓同一套基因組在不同環境訊號下展現不同結果。當代更精確的提問方式是:基因與環境如何在發育的每一個時間點交互作用(gene-environment interaction)?例如同一個風險基因,在高壓力與低壓力環境下對發展軌跡的影響可能截然不同。
第二,「關鍵期」是否真的不可逆,正在被重新檢視。 經典觀點認為錯過關鍵期就無法挽回,但近年研究發現關鍵期的「開」與「關」本身是由分子煞車(如抑制性 GABA 神經元的成熟、神經周圍網 PNN 的形成)主動控制的。這意味著理論上有可能透過藥理或行為手段「重新開啟」可塑性——這對中風復健、弱視治療乃至成人學習都有深遠意涵。這也讓「終身學習」有了更紮實的神經科學基礎:成人腦的可塑性雖不如幼年,但遠非定型。
第三,發育觀點正在重塑我們對精神疾病的理解。 思覺失調症(schizophrenia)、自閉症光譜(autism spectrum)等狀況愈來愈被視為「神經發育疾病」(neurodevelopmental disorders)——問題可能源於早期遷徙、突觸形成或修剪的細微偏差,卻要到多年後才以症狀顯現。有一個影響深遠的假說認為,思覺失調症的部分成因是青春期「突觸修剪過度」,這把研究的時間焦點從成年往前推到發育早期。
最後,神經發育與人工智慧(AI)的對話正在加深。 人工神經網路的訓練,某種意義上與大腦的「修剪」異曲同工:深度學習中的「剪枝」(pruning)技術刻意移除冗餘參數以提升效率,靈感正部分來自突觸修剪;而「關鍵期」現象在人工網路訓練中也被觀察到——模型在訓練早期接收到的資料分布,會對最終能力產生不成比例的影響。反過來,AI 模型也成為研究大腦發育的工具:研究者用計算模型模擬皮質分層、軸突尋路與表徵學習的湧現過程。這種雙向啟發提醒我們,無論是碳基還是矽基的智能,「如何從簡單規則長出複雜結構」都是同一個根本問題。對於在 Uedu 上跨領域學習的同學而言,神經發育恰好站在生物、心理與計算科學的交會點,是理解「智能如何成形」最具啟發性的入口之一。