
同樣是負回饋,為什麼有些變數能「精準歸零」、有些只能「大致拉回」?
用控制理論拆開恆定:比例控制的殘差、積分控制的完全適應、增益與延遲造成的震盪,以及前饋與分子層次的實作。
同樣是負回饋,為什麼有些變數能「精準歸零」、有些只能「大致拉回」?
入門篇告訴我們,身體靠負回饋(negative feedback)把偏離設定點的變數拉回去。但若你仔細追問,會發現一個耐人尋味的不對稱:有些生理變數在擾動過後能幾乎完全回到原值,誤差近乎為零;有些變數卻只能被「大致」拉回,永遠殘留一點偏移。
舉個對照。一個健康的人長期每天多吃幾克鹽,他的細胞外液量(extracellular fluid volume)與動脈血壓,在數天之後幾乎會回到原本的水準——彷彿身體擁有一個能「記住原點」的會計系統。可是同一個人若搬到高海拔,動脈血氧分壓(PaO₂)即使經過數週適應,仍會停在低於海平面的數值,只是被部分代償。為什麼同樣標榜負回饋,調節的「徹底程度」差這麼多?
答案藏在控制理論(control theory)裡:負回饋只是一個大類,其內部還分成比例控制(proportional control)與積分控制(integral control),兩者在數學上有本質差異。 入門篇把負回饋當成一個整體概念;進階篇要把它拆開,看清楚生理系統如何用不同的控制策略,換取不同的精度、速度與穩定性。理解這層差異,你才會明白為什麼血壓的長期調控非靠腎臟不可,也才會明白為什麼有些代償永遠補不滿。
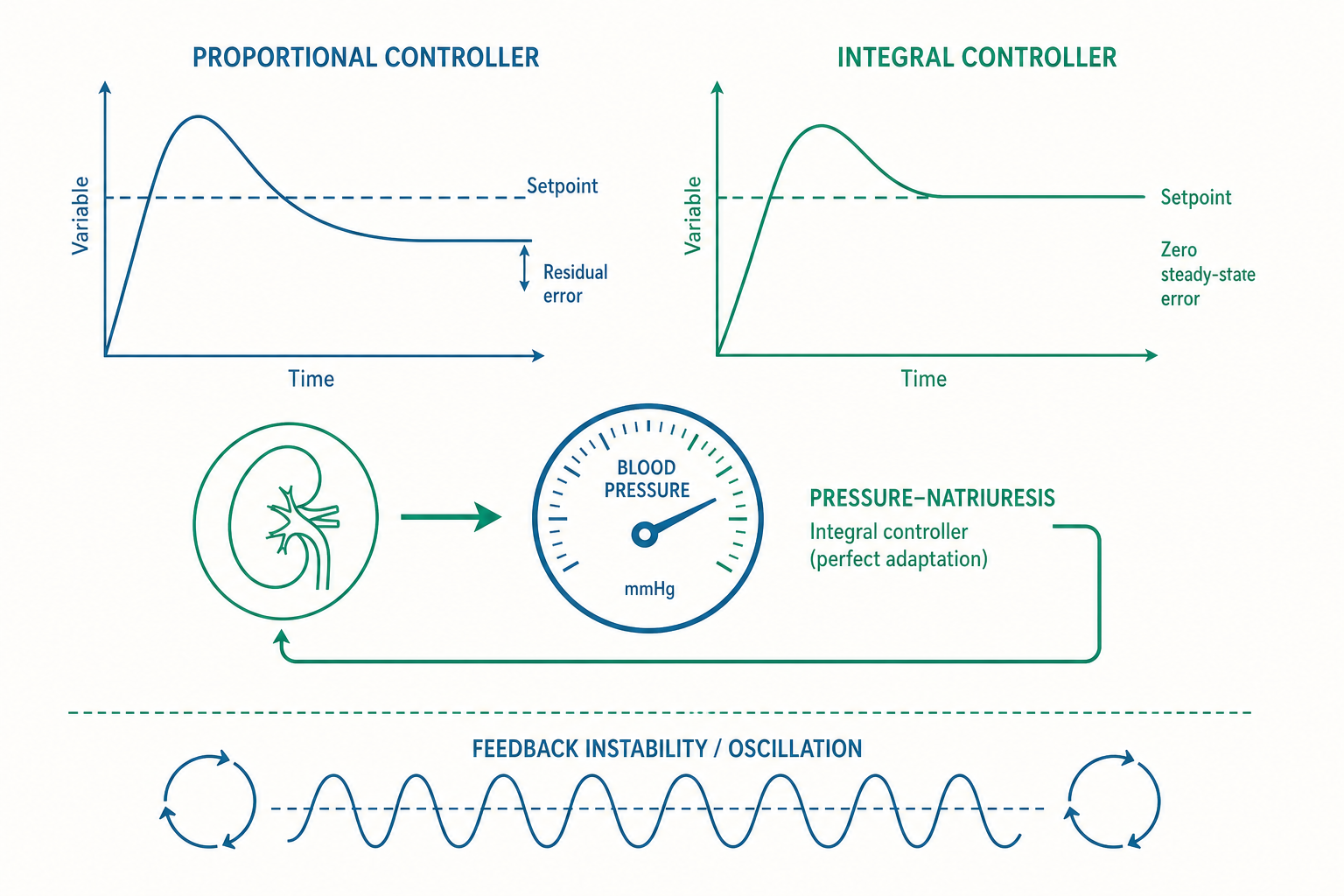
比例控制與它無法擺脫的殘差
先看最常見的一類負回饋:比例控制。它的核心是「反應強度正比於誤差大小」。誤差越大,效應器出力越猛;誤差縮小,出力跟著減弱。許多快速生理反射屬於這一型,例如壓力感受器(baroreceptor)反射對血壓的瞬間調節。
比例控制反應快、結構簡單,但它有一個無法迴避的數學宿命:穩態殘差(steady-state error)。原因可以用一句話講清楚——只要誤差被縮小到零,效應器的「驅動力」也跟著歸零;可是若擾動仍然存在,沒有驅動力就守不住,於是系統只能停在一個「誤差不為零、但剛好小到讓殘餘驅動力能對抗擾動」的折衷點。換句話說,比例控制天生需要保留一點誤差,才能維持讓它工作的動力。
把它寫成最精簡的關係:若效應器輸出 = 增益(gain, G)× 誤差,而要對抗的擾動需要某個固定輸出,則穩態時必然殘留 誤差 = 擾動 ÷ G。增益越大,殘差越小,但不會真正等於零;而且增益若一味調高,又會引發過衝與震盪(後面會談)。這就是為什麼純比例的反射,例如急性壓力感受器反射,能在你站起來的瞬間把血壓「大致」撐住,卻不擅長把血壓「精確」固定在某個長期目標——它本來就不是為長線歸零而設計的。
積分控制:身體如何達成「robust perfect adaptation」
那麼,那些能精準歸零的變數,靠的是什麼?答案是積分控制:效應器的輸出不只取決於當下的誤差,而是取決於誤差隨時間累積的總和(積分)。
積分控制有一個漂亮的性質。只要誤差不等於零,積分項就會持續累積、輸出持續變化,系統絕不肯停下來——唯有當誤差「恰好等於零」時,積分才不再增減、輸出才穩定。換言之,積分控制的穩態解強迫誤差為零,而且這個歸零不依賴增益的大小、也不依賴擾動的大小。這種「不論擾動多大、最終都回到同一原值」的能力,系統生物學稱為穩健完全適應(robust perfect adaptation, RPA)。
生理學中最具代表性的積分控制器,是腎臟對細胞外液量與血壓的長期調節,也就是 Arthur Guyton 提出的壓力—利鈉(pressure-natriuresis)機制。它的邏輯是:動脈壓一旦高於某個目標,腎臟排出的鈉與水就增加;只要血壓還高於目標,腎臟就「一直排、一直排」,體液量持續下降、血壓持續往下走,這個排泄不會因為血壓只高一點點就停手——它只在血壓回到目標點時才平衡。正因為腎臟把「血壓誤差」對時間做了積分,長期血壓設定點才能被守得近乎絕對。Guyton 由此提出一個在循環生理學中相當激進的論點:長期動脈血壓的設定,最終由腎臟的鹽水平衡曲線決定,而不是由血管阻力或心輸出量單獨決定。 這也是為什麼幾乎所有有效的長期降血壓策略,最終都要動到腎臟處理鈉的能力。
對照之下,缺氧的代償為什麼補不滿?因為頸動脈體(carotid body)的化學感受器對低血氧的反應比較接近比例型,加上紅血球生成、通氣等代償各有上限與副作用(過度通氣會洗掉 CO₂ 造成鹼中毒),系統最終停在一個帶殘差的折衷,而非真正的零誤差適應。同樣是負回饋,控制器的型態(比例 vs 積分)決定了它究竟能「歸零」還是只能「逼近」。
看一個例子:高鹽飲食數天後,血壓為何回得來?
讓我們把積分控制套到一個具體情境,追蹤時間軸。
一位健康受試者連續一週每天多攝取約 10 克鹽。第一天,鈉與水滯留,細胞外液量增加,血壓略升——比例型的快速反射(壓力感受器)會先讓血管放鬆、心跳放慢,把急性的升幅壓下一部分,但這只是把「斜率」磨平,沒有解決根本的鈉超載。
接著腎臟的積分控制登場。只要血壓仍高於原本的目標,壓力—利鈉效應就讓腎臟持續多排鈉與水。注意「持續」這個詞:哪怕血壓只比目標高 2 mmHg,腎臟也不會滿足於現狀,而是日復一日地把多餘的鈉排掉,直到血壓回到原點、利鈉的驅動才消失。於是在數天之後,儘管每天攝取的鹽依舊偏高,血壓卻已悄悄回到接近基線——這正是 RPA 的臨床表現:對「持續存在的擾動」達成完全適應。
這個例子也揭示了一個臨床洞見:在某些「鹽敏感型高血壓(salt-sensitive hypertension)」病人身上,腎臟的壓力—利鈉曲線發生了右移或變平,等於把這個積分控制器的「目標點」調高了。於是同樣的鹽負荷,他們得用更高的血壓才能把鈉排乾淨——身體不是壞了負回饋,而是把積分控制器歸零的那個原點,整個往上搬了。理解這層,就能看懂為什麼限鹽與利尿劑對這群病人特別有效:它們都是在替那條失準的鹽水曲線「補課」。
增益、時間延遲與震盪:負回饋的暗黑面
入門篇把負回饋描繪成穩定的守護者,但控制理論提醒我們:負回饋若設計不當,反而會自己製造不穩定。 關鍵在三個量的互動:增益(gain)、時間延遲(time delay)與系統的慣性。
直覺上你可能以為增益越高、調節越好——畢竟前面說了高增益能壓低比例控制的殘差。但增益太高加上回饋有時間延遲,會釀成過衝(overshoot)與震盪(oscillation):感受器測到的是「過去」的狀態,效應器卻在「現在」用力修正,等修正生效時情況早已改變,結果一下子矯枉過正、一下子又補過頭,變數開始來回擺盪。工程上把這種因延遲而生的不穩定用相位邊界(phase margin)來描述,而生理系統裡到處可見它的影子。
最經典的臨床例子是陳—施呼吸(Cheyne–Stokes respiration):呼吸由深快逐漸轉為微弱、甚至停止(呼吸暫停),再由弱轉強,周而復始。它的本質就是化學感受器回饋環的震盪——當心衰竭使血液從肺循環到腦部化學感受器的「循環時間(circulation time)」延長(延遲變大),或感受器對 CO₂ 的敏感度(增益)異常增高,這個負回饋系統就跨過了穩定邊界,從「平穩調節」滑向「自發震盪」。換句話說,陳—施呼吸不是隨機故障,而是一個增益過高、延遲過長的負回饋系統可預測的數學行為。
這帶出一個深刻的觀念轉換:許多生理性與病理性的節律(呼吸的、荷爾蒙脈衝式分泌的、甚至某些心律不整的),都可以被理解為負回饋環在特定參數下「不再收斂、而是進入極限環(limit cycle)」的結果。穩定與震盪,往往只隔著一個增益或一段延遲。
前饋控制:在擾動到達之前就先出手
到目前為止談的都是回饋(feedback)——等變數先偏離、感受器測到、再修正。但身體還有一套更聰明的策略:前饋控制(feedforward control),在擾動真正造成偏離「之前」就預先反應。
回饋的根本弱點是它必然「慢半拍」:一定要先產生誤差,才有東西可修。前饋則繞過這個瓶頸——它根據「即將到來的擾動」本身,預先啟動代償。最清楚的例子是進食的頭期反應(cephalic phase):你光是看到、聞到、咀嚼食物,副交感神經就已經命令胃分泌胃酸、胰臟分泌少量胰島素,搶在葡萄糖湧入血液之前先做好準備。又如運動:在肌肉真正缺氧、CO₂ 真正堆積之前,大腦運動皮質的「中樞命令(central command)」就同步調高心率與通氣——這就是為什麼你一起跑,心跳幾乎是瞬間就上去,而不是等到乳酸堆積才慢慢爬升。
成熟的生理調節幾乎都是前饋+回饋的混合:前饋負責快、負責預判,但它「盲目」——若預測錯了沒有自我校正能力;回饋負責準、負責收尾,把前饋沒算對的殘差補平。理解這個分工,你會對「為什麼身體既快又準」有更立體的答案:不是單靠某一種控制,而是兩種互補策略的協作。
動手試試:用一個思想實驗分辨控制器型態
你不需要任何儀器,只要拿幾個生理現象來分類,就能訓練自己用控制理論的眼光看身體。對下列每個反應,問自己三個問題:(1) 它反應快還是慢?(2) 擾動過後變數能完全歸零,還是殘留偏移?(3) 它是等誤差出現才動(回饋),還是預先動(前饋)?
- 站起來時心跳瞬間加快:快、回饋、比例型——它撐住瞬間血壓,但不負責長期血壓的精確設定。
- 長期高鹽後血壓數天回到基線:慢、回饋、積分型——靠腎臟壓力—利鈉達成近乎完全的適應。
- 看到食物就分泌唾液與胰島素:前饋——擾動還沒到就先反應,沒有誤差也照樣啟動。
- 發抖產熱對抗寒冷:回饋、比例型——冷得越厲害抖得越猛,但體溫通常仍略低於設定點,殘差存在。
做完這個分類,你會發現一件事:入門篇所說的「負回饋」其實是一個內部結構豐富的家族,而把每個反應放進「快/慢 × 比例/積分 × 回饋/前饋」這張坐標,正是進階生理學分析任何調節迴路的起手式。
分子層次:細胞如何「硬體實作」一個積分控制器
控制理論講得再漂亮,最終仍要問:細胞用什麼分子機器把積分控制蓋出來?系統生物學近二十年的一大進展,就是找到了能達成穩健完全適應(RPA)的最小分子拓樸。
研究發現,要在分子網路裡實現完全適應,幾乎都繞不開一個關鍵母題:抗衡的整合(antithetic integral) 或 不連貯前饋環(incoherent feedforward loop)。前者由一對互相中和的分子(例如一個會「抓住」並抵消另一個的分子對)構成,其數學行為恰好等於對誤差做積分——這正是工程積分器的生化版本。細菌趨化(bacterial chemotaxis)系統就是最早被嚴格證明具有 RPA 的例子:受體甲基化與去甲基化的酵素活性差,扮演了積分器的角色,使細菌無論背景化學濃度多高,都能對「濃度的相對變化」保持一致的靈敏度。
這個發現的意義超越微生物學。它告訴我們,「完全適應」不是某個器官的特異功能,而是一種可以被通用分子拓樸實現的設計原則;從細菌的趨化、到鈣離子恆定中副甲狀腺—腎—骨的迴路、再到血壓的長期調控,自然界一再用相似的網路骨架達成同一件事。對醫學生而言,這提供了一條把分子生物學、系統生理學與臨床現象串起來的主軸:當你看到一個變數能「完全適應」,就該去找實作它的那個積分母題;當適應失準,問題往往出在那個母題的某個參數被改寫了。
重點回顧
- 負回饋不是單一機制,而是包含比例控制與積分控制等不同型態的家族;兩者數學性質不同,導致調節的徹底程度截然不同。
- 比例控制反應快但有無法消除的穩態殘差:誤差歸零則驅動力歸零,故必須保留誤差才守得住;高增益能縮小殘差卻可能引發震盪。
- 積分控制對誤差隨時間累積,穩態必然強迫誤差為零,達成穩健完全適應(RPA);腎臟的壓力—利鈉是長期血壓調控的積分器,故 Guyton 主張長期血壓最終由腎臟鹽水平衡決定。
- 增益過高+時間延遲會讓負回饋從穩定滑向震盪(極限環);陳—施呼吸正是化學感受器回饋環因循環時間延長而自發震盪的臨床範例。
- 前饋控制在擾動到達前預先反應(頭期反應、運動中樞命令),與回饋互補:前饋快但盲目、回饋準但慢半拍;成熟調節幾乎都是兩者混合。分子層次上,RPA 由抗衡積分/不連貫前饋等通用拓樸實作。
深入探討(研究所視角)
走到研究前沿,這套「把生理當控制系統」的視角會展開成好幾條值得深耕的路線。
RPA 的拓樸普適性與限制。 近年系統生物學以嚴格數學證明:能在生化網路中達成穩健完全適應的拓樸其實相當有限,可被歸納成少數幾類核心母題。這個結果由 Araujo 與 Liotta 等人推進,把「哪些網路結構必然能完全適應」變成可窮舉的設計問題。對生理學的啟示是:當我們在不同器官、不同物種反覆看到同一種積分式調控,未必是巧合,而可能是「能達成此功能的結構本就不多」的數學必然。值得思考的延伸是——若某條調控環演化出 RPA,它在何種擾動下會失效?因為 RPA 的「穩健」通常只對某一類擾動成立,對另一類(例如改變積分器本身參數的擾動)反而毫無抵抗力,這正對應許多疾病「悄悄移動設定點」的隱匿性。
設定點究竟是不是真的存在? 入門篇與本文都假設有個「設定點」讓系統去歸零,但這個概念在學界其實有爭議。積分控制的數學告訴我們:一個系統可以表現得「彷彿有設定點」,卻不需要在任何地方真的儲存一個目標數值——設定點可以是網路動力學的湧現性質(emergent property),而非某個被寫死的參數。這個觀點(有時稱為 settling point 而非 set point)對體重調節、體溫調節的研究影響深遠:它解釋了為什麼減重後身體會頑強反彈(積分器把你拉回它隱含的平衡),也提醒我們別天真地以為「調高某個旋鈕」就能重設一個其實是分散式湧現出來的目標。
從個體到計算模型:in silico 生理學。 Guyton 在 1972 年就提出了一個包含數百個方程式、涵蓋循環—腎臟—體液—激素的整合模型,是計算生理學的里程碑。今天,這類大型模型已用於藥物的心臟安全評估、敗血症與重症的虛擬病人模擬,乃至個人化醫療。它們的共同精神是:單一器官的直覺往往會誤導,唯有把多個回饋與前饋環一起解,才能預測跨系統的非直覺行為(例如為何單純擴張血管的藥物,長期下來血壓常被腎臟的積分控制「追平」而失效)。
對教育科技的跨域連結。 把學習者視為一個需要被調節的動態系統,這套控制語言同樣適用:認知負荷、注意力、壓力都有各自的「快速比例反射」與「慢速積分漂移」。Uedu 的 Educational Omics 框架中,PhysioNeuromics 維度蒐集 HRV、睡眠、壓力等時序資料,而 HRV 本身就是自律神經回饋環增益與延遲的外顯指標;Uedu Mind 的 PALM(生理感知語言模型)試圖整合這些訊號以判讀學習狀態,背後的理論骨架,與本文描述的生理控制系統一脈相承。把恆定的數學語言帶進學習分析,或許正是「生理學 × 資料科學 × 教育」這個交叉口最值得探索的方向。
把「負回饋」拆成比例與積分、再補上前饋與分子實作,你手上就不再只有一個籠統的恆定概念,而是一套能定量分析、能預測震盪、能定位疾病、甚至能跨界到學習科學的分析工具。這正是從入門邁向進階的關鍵一步:不只知道身體「會」維持穩定,而是能說清楚它「用哪一種控制策略、付出什麼代價、在哪個參數失準時就會生病」。
(本文為醫學教育性內容,旨在以控制理論視角深化對生理調節機制的理解,不構成個人醫療診斷或治療建議。若有健康疑慮,請諮詢合格醫療專業人員。)