
一場為期數百萬年的軍備競賽,藏在你的淋巴結裡
從生發中心的體內天擇、抗體類型轉換,到病原體逃脫術與抗藥性演化,看懂宿主與微生物的共演化軍備競賽。
一場為期數百萬年的軍備競賽,藏在你的淋巴結裡
在入門篇裡,我們把感染想像成一道傷口背後的攻防戰:病原體入侵、先天免疫搶先反應、適應性免疫帶著記憶精準收尾。那是一張清楚的「組織圖」。但如果你已經熟悉這張圖,接下來真正有趣的問題會是:這套系統是怎麼「演化」出來的?而病原體又怎麼「反演化」回去?
換句話說,免疫不是一套固定的劇本,而是一場持續進行、雙方都在即時更新策略的共演化軍備競賽(coevolutionary arms race)。這場競賽有兩個時間尺度:一個是百萬年的演化尺度,宿主與病原體互相塑造彼此的基因組;另一個是短短幾天到幾週的個體尺度——你的免疫系統會在一次感染中,於淋巴結裡進行一場微縮版的「達爾文演化」,當場篩選出更厲害的抗體。這篇進階文章,要帶你深入這場競賽的分子細節:抗體如何在體內被「演化」得越來越精準、病原體用哪些招數逃脫、以及細菌的抗藥性如何在數小時內擴散。
(提醒:本文為醫學知識讀本,旨在說明生理與病理機制及其研究意涵,不構成個人醫療建議。任何健康疑慮請諮詢專業醫師。)
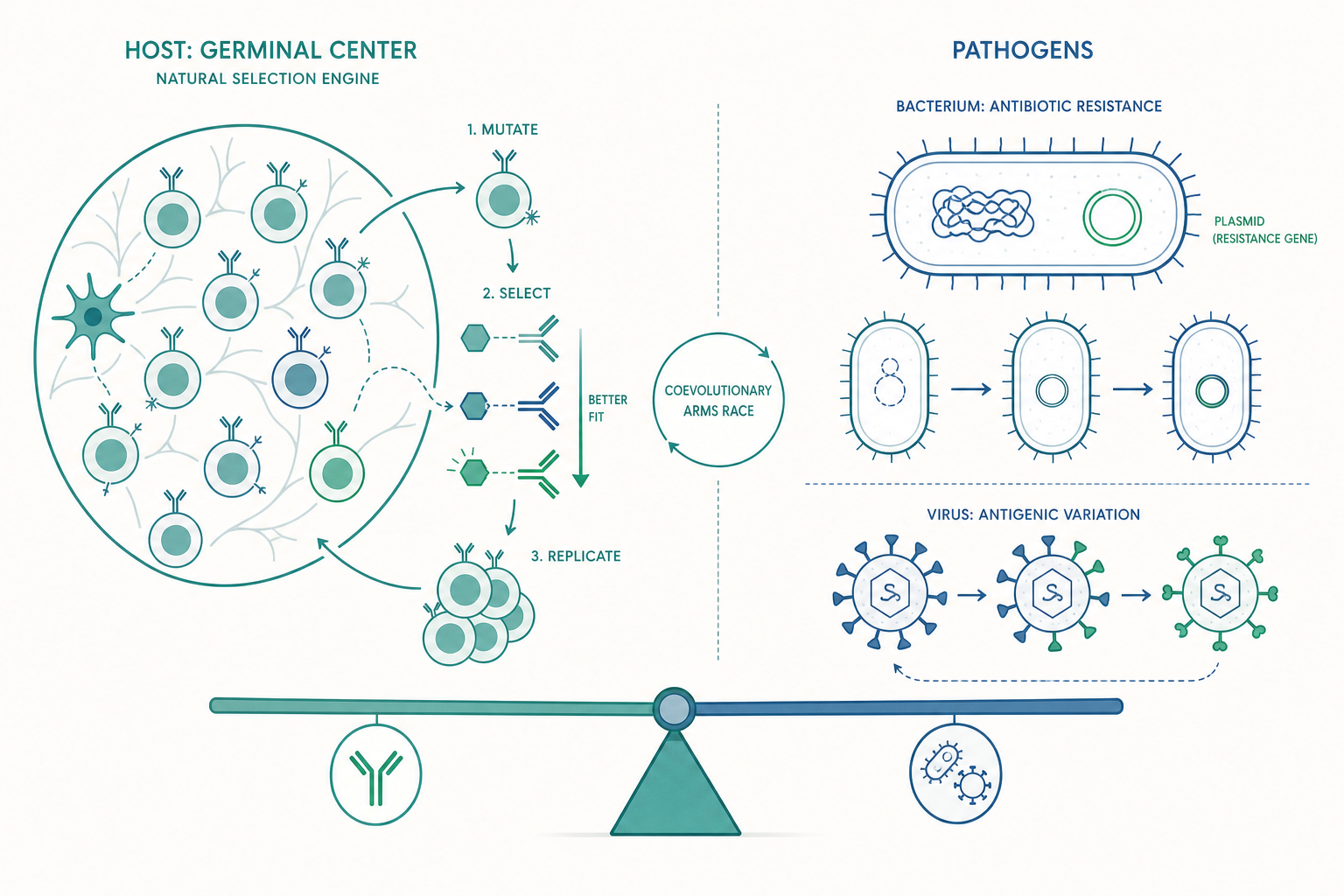
淋巴結裡的達爾文:親和力成熟與生發中心
入門篇提過 B 細胞會產生抗體、留下記憶細胞。但有個問題被略過了:初次感染時冒出來的抗體,其實品質相當粗糙。 它們能「大致認得」抗原,但結合得不夠緊。免疫系統真正厲害之處,是它能在感染過程中,把這些平庸的抗體「升級」成結合力強上千倍的精品。這個過程稱為親和力成熟(affinity maturation),發生的場所是淋巴結裡的生發中心(germinal center, GC)。
生發中心本質上是一座微型的「天擇引擎」,它把達爾文演化的三要素——變異、選擇、繁衍——壓縮在幾天內反覆運轉:
-
變異(突變):被活化的 B 細胞會啟動一種叫體細胞超突變(somatic hypermutation, SHM)的機制。一個關鍵酵素 AID(activation-induced cytidine deaminase,活化誘導胞苷脫胺酶)會專門在抗體基因的可變區引入點突變,突變率高達正常基因的百萬倍。注意這是「刻意」把錯誤集中在抗體基因上——免疫系統允許自己在這一小段 DNA 上「亂寫」,藉此產生大量結合力各異的變異抗體。
-
選擇:突變後的 B 細胞必須去爭奪有限的抗原。生發中心裡的濾泡樹突細胞(FDC)表面展示著抗原,B 細胞用自己新突變出的受體去「搶」抗原。結合得越緊的 B 細胞,搶到越多抗原,再把抗原呈現給濾泡輔助型 T 細胞(Tfh)以換取存活訊號。結合力弱的 B 細胞搶不到訊號,就地凋亡淘汰。
-
繁衍:勝出的 B 細胞被送回去再分裂、再突變,進入下一輪循環。
經過數輪「突變—競爭—淘汰」,抗體對抗原的親和力(affinity)可以提升 10 到 1000 倍。這就是為什麼初次感染後幾週、以及打完疫苗一段時間後,抗體品質才達到高峰——你的身體正在淋巴結裡跑一場小型的天擇實驗。
理解這一點有深遠意義:疫苗追加劑(booster)的時機,本質上是在調控這場演化的節奏。 間隔太短,記憶 B 細胞還來不及完成親和力成熟;適當延長間隔,反而能讓抗體品質更好——這是疫苗學裡「prime-boost」策略背後的免疫學原理。
抗體類型轉換:同一支抗體,換上不同的「戰鬥服」
親和力成熟調的是抗體的「辨識端」(可變區)。但抗體還有另一端——恆定區(constant region),決定它的「功能性格」。同一個 B 細胞選殖株(clone)在演化出高親和力受體的同時,還能進行類型轉換重組(class switch recombination, CSR),把抗體的恆定區從 IgM 換成 IgG、IgA 或 IgE,而辨識端維持不變。
這就像同一名特務,辨識目標的本事沒變,但能依任務換不同裝備:
- IgM:初次感染最早出現,五聚體結構、活化補體能力強,但親和力較低、壽命短。臨床上驗到病原體特異性 IgM,常代表「近期或正在感染」。
- IgG:血液中最主要、壽命最長的抗體,是親和力成熟後的主力,能穿過胎盤把母體免疫力傳給胎兒。驗到特異性 IgG 常代表「曾經感染或已建立記憶」。
- IgA:黏膜(腸道、呼吸道、母乳)的守門員,以雙聚體形式分泌到體表黏液中,是第一線阻擋病原體附著的關鍵。
- IgE:濃度極低,但綁在肥大細胞上負責對抗寄生蟲——也是引發過敏反應的元兇。
哪一種類型被選擇,取決於 Tfh 細胞分泌的細胞激素(cytokine)環境。這帶出一個關鍵概念:免疫反應的「方向」是被調控出來的,不是預設的。 同一個抗原,在不同的細胞激素環境下,會被導向截然不同的反應類型——這也是為什麼某些疫苗佐劑(adjuvant)的設計重點,是去「形塑」免疫反應的型別,而不只是放大它。
看一個例子:流感病毒為何能年年捲土重來
入門篇提過流感疫苗要每年重打,因為病毒抗原會變。進階一點看,這背後是兩種截然不同的演化機制,理解它們能讓你看懂大流行(pandemic)與季節性流感的差別。
抗原漂移(antigenic drift)——慢慢偷改。 流感病毒的 RNA 聚合酶沒有校對功能,複製時錯誤率極高。在人群免疫力的壓力下,那些剛好突變到「現有抗體認不太出來」的病毒株會被天擇選出來,逐年累積。這是一種漸進的逃脫,造成季節性流感,也是疫苗需年年更新的原因。
抗原移型(antigenic shift)——整段換掉。 流感病毒基因組分成 8 段。當兩種不同來源的流感病毒(例如人流感與禽流感)同時感染一個宿主細胞(豬常被視為「混合容器」),它們的基因段可以重新洗牌(reassortment),組合出一個帶有人群從未見過的全新表面抗原的病毒。由於幾乎沒人有免疫記憶,這種劇變正是引發全球大流行的危險來源,1918、1957、2009 年的大流行都與此有關。
用一句話總結這場例子的教訓:病原體的演化速度,決定了我們防禦策略的有效期限。 漂移讓我們疲於追趕,移型則可能讓既有防線一夜失效。這也是為什麼開發「廣效流感疫苗」(針對病毒較不易突變的保守區段)是當代疫苗學的聖杯之一。
病原體的逃脫術:當對手也會演化
宿主有生發中心這樣的演化引擎,病原體當然不會坐以待斃。它們發展出一整套對抗免疫系統的策略,每一種都針對前面提過的某個防禦環節:
-
抗原變異(antigenic variation):除了流感的漂移與移型,非洲錐蟲(引起昏睡病)更誇張——它有上千個表面醣蛋白基因,能不斷輪換表面外衣,等抗體好不容易追上,它已經換了一身。瘧原蟲也用類似策略。
-
躲進細胞、關掉展示窗:入門篇提過 CD8⁺ T 細胞靠 MHC class I 這扇「展示窗」看見細胞內的病毒。某些病毒(如巨細胞病毒 CMV、某些皰疹病毒)會主動下調宿主細胞的 MHC class I 表現,等於把窗簾拉上、躲過 T 細胞巡查。但宿主也有對策:自然殺手細胞(NK cell)專盯「不展示 MHC 的可疑細胞」,這就是有名的「失蹤的自我(missing self)」假說。攻防雙方環環相扣。
-
干擾補體與吞噬:許多細菌長出多醣莢膜(capsule),讓補體與吞噬細胞抓不住自己(肺炎鏈球菌就是經典例子,這也是莢膜疫苗的標靶)。
-
潛伏(latency):皰疹病毒、結核桿菌、HIV 都會進入潛伏狀態,把基因藏在宿主細胞裡幾乎不表現,免疫系統便「看不到」它。等宿主免疫力下降時再重新活化。這正是為什麼這些感染極難根治——它不是被殺光,而是「藏起來」了。
把這些放在一起看,會浮現一個深刻的圖像:免疫系統的每一道防線,都對應著病原體的一套反制手段,而每套反制又催生宿主的新對策。 這場競賽沒有終點,只有不斷升級的平衡。紅皇后假說(Red Queen hypothesis)形容得貼切:雙方都得拚命奔跑,才能停在原地。
抗藥性:一場我們正在加速的演化
如果說免疫與病原體的軍備競賽是自然演化的產物,那麼抗生素抗藥性(antibiotic resistance)就是人類正在親手加速的一場演化災難,也是當代公共衛生最嚴峻的挑戰之一。
抗生素的選擇壓力極其直白:施加抗生素,沒有抗藥性的細菌死光,剛好帶有抗藥基因的少數細菌活下來、大量繁殖。抗生素不會「製造」抗藥性,它只是強力篩選出本來就存在的抗藥變異。 細菌獲得抗藥能力的機制大致有四類:
- 酵素分解或修飾藥物:如 β-內醯胺酶(β-lactamase)能水解青黴素的核心結構,使藥物失效。
- 改變藥物標靶:MRSA(抗甲氧西林金黃色葡萄球菌)改造了青黴素結合蛋白,讓藥物抓不到目標。
- 降低藥物進入或加速排出:改變細胞膜通透性,或用外排幫浦(efflux pump)把藥物泵出去。
- 繞道(bypass):發展出不被藥物影響的替代代謝途徑。
更棘手的是,抗藥基因常坐落在質體(plasmid)這種可獨立複製的環狀 DNA 上,而細菌能透過水平基因轉移(horizontal gene transfer)——尤其是接合作用(conjugation)——把整個質體傳給鄰居,甚至傳給不同菌種。這意味著抗藥性可以橫向擴散,速度遠快於垂直遺傳。一個帶有多重抗藥質體的腸道細菌,可能在數小時內就把抗藥武器分發給整個菌群。
動手試試:用演化邏輯推理臨床指引
不妨用這套演化思維,重新檢視幾個你或許聽過、卻未必想透原理的臨床原則:
- 為什麼醫師再三叮嚀「抗生素要吃完整個療程」(在傳統指引下)?
- 為什麼農畜業大量在飼料裡添加抗生素,會被公共衛生界強烈反對?
- 為什麼「組合用藥」(同時用多種抗生素)能延緩抗藥性出現,常用於結核病治療?
參考思路:(1) 提前停藥時,藥物剛好只清掉最敏感的菌、留下耐受性較高的「半抗藥」族群繼續繁殖,等於替抗藥性鋪路(補充:近年部分研究對「一律吃完」提出更細緻的修正,強調應依感染類型與證據個別化,但「不可隨意亂停、亂用」的核心不變)。(2) 飼料抗生素等於對全國牲畜的菌群長期施加微量選擇壓力,是培養抗藥菌的溫床,且可透過食物鏈與環境傳給人。(3) 細菌要同時演化出對多種藥物的抗性,機率是各別機率的乘積,遠低於對單一藥物——多管齊下讓它「無路可逃」。結核病療程要合併數種藥物、持續數月,正是基於此邏輯。
能用「選擇壓力」與「機率乘積」這兩個概念把三題串起來,代表你已經能把演化生物學直接應用到臨床決策上了——這正是進階學習者該有的視角。
量化的免疫學:用數字看懂這場戰爭
進階學習的另一個面向,是把免疫從「故事」推向「數量」。臨床與研究越來越倚賴量化指標來描述這場攻防,舉幾個有代表性的例子:
-
抗體效價(titer)與中和抗體:驗血報告上的抗體效價,是把血清連續稀釋到剛好還測得到抗體的倍數。但「測得到抗體」不等於「有保護力」——真正關鍵的是中和抗體(neutralizing antibody),也就是能實際阻止病毒入侵細胞的那部分。一個重要的研究概念是保護相關指標(correlate of protection):找出「抗體達到多少效價,就有幾成保護力」的量化關係,這是疫苗評估的核心。
-
病毒量與檢測極限:COVID-19 期間普及的 PCR 檢測會給出 Ct 值(循環閾值)——病毒量越高,越少的擴增循環就能驗到訊號,Ct 值越低。理解 Ct 值是對數尺度,能幫你正確解讀「Ct 從 20 升到 30 代表病毒量約掉了 1000 倍」這類資訊,而不是線性地以為「只少了一半」。
-
基本再生數 R₀ 與免疫學的連結:流行病學的 R₀(一個感染者平均傳染給幾人)看似宏觀,其實深植於個體免疫。群體免疫門檻約為 1 − 1/R₀:R₀ 越大,需要越高比例的人有免疫力才能阻斷傳播。麻疹 R₀ 高達 12–18,所以需要約 95% 接種率;這個數字直接決定了公共衛生政策。
這些量化工具的共同價值在於:它們讓免疫從定性的「強/弱」描述,變成可比較、可預測、可決策的科學。 進階學習者應習慣於在讀到任何免疫數據時,先問三件事:這個數字是線性還是對數尺度?它測的是「有沒有」還是「夠不夠」?它和臨床保護力的關係被驗證過嗎?
重點回顧
- 生發中心是體內的天擇引擎:透過體細胞超突變(AID 酵素)製造變異、靠搶奪抗原進行選擇,讓抗體在數天內完成親和力成熟,結合力提升可達千倍——疫苗追加劑的時機本質上在調控這場演化。
- 類型轉換(CSR)讓同一支抗體換功能:辨識端不變、恆定區改換(IgM/IgG/IgA/IgE),由細胞激素環境決定方向;免疫反應的「型別」是被調控出來的。
- 病原體有整套逃脫術:抗原漂移與移型、下調 MHC、莢膜抗吞噬、潛伏藏匿——每一招都對應宿主的某道防線,催生紅皇后式的持續軍備競賽。
- 抗藥性是人為加速的演化:抗生素只是篩選而非製造抗藥性;質體與水平基因轉移讓抗藥基因能跨菌種橫向擴散,組合用藥靠「機率乘積」延緩抗性。
- 量化視角是進階關鍵:分辨中和抗體與一般抗體、看懂 Ct 值的對數尺度、理解 R₀ 與群體免疫門檻的連結,才能正確解讀免疫數據。
深入探討(研究所視角)
對希望進一步鑽研的學習者,以下幾條線索通往當代免疫學與微生物學最前沿、也最具跨領域張力的議題。
生發中心動力學的單細胞與活體影像研究。 親和力成熟過去只能從結果推測,如今雙光子活體顯微鏡(intravital two-photon microscopy)已能即時觀察 B 細胞在生發中心明區與暗區之間往返的動態,而單細胞定序(single-cell sequencing)則能重建一個 B 細胞選殖株在感染過程中的完整突變譜系樹(lineage tree),等於把演化過程「拍成影片」。這讓免疫學進入可定量建模的階段:研究者用演化動力學方程式去預測,怎樣的抗原暴露序列能引導抗體往「廣效中和」方向演化——這正是對抗 HIV、流感等高變異病原體之疫苗設計的核心難題。
結構疫苗學(structural vaccinology)與抗原工程。 既然抗體與抗原的結合本質是分子結構的契合,當代疫苗設計已從「拿整隻病原體」進化到「以原子層級設計抗原」。研究者用冷凍電子顯微鏡(cryo-EM)解析病毒表面蛋白與中和抗體結合的精確構象,再反向設計出穩定在最易被中和構象的抗原(如將呼吸道融合病毒 RSV 的 F 蛋白鎖在融合前構象),大幅提升疫苗效力。AI 蛋白質結構預測(如 AlphaFold)正加速這個領域,使「先在電腦裡設計抗原、再合成驗證」成為可能。
抗藥性的演化軌跡可預測性與精準抗生素策略。 抗藥性研究的前沿問題是:細菌走向抗藥的演化路徑是否可預測、可預先封堵?研究者用實驗演化(experimental evolution)配合深度定序,繪製抗藥性突變的適應度地形(fitness landscape),發現某些路徑存在「演化捷徑」與「死路」。由此衍生出附帶敏感性(collateral sensitivity)策略——找出「對 A 藥產生抗性,反而會對 B 藥變敏感」的權衡關係,再設計藥物輪替方案,引誘細菌走進演化死巷。這把演化生物學、基因體學與臨床藥理學緊密縫合在一起。
噬菌體療法與 CRISPR:把微生物的武器轉為己用。 面對抗藥性危機,一條復興中的路線是噬菌體療法(phage therapy)——用專門感染細菌的病毒去精準獵殺特定致病菌。其精妙在於噬菌體與細菌本身就在共演化:細菌演化出抗噬菌體機制(CRISPR-Cas 系統最初正是細菌的「抗病毒免疫」),人類則反過來把 CRISPR 改造成基因編輯工具。這條從「細菌免疫系統」到「人類基因編輯技術」的轉化路徑,是基礎微生物學意外結出果實的典範,也提醒我們:微生物世界的攻防智慧,往往遠早於、且遠精巧於人類的設計。
跨領域連結:免疫的演化—生態—系統思維。 把以上線索拉高來看,會發現當代免疫學正在從「分子機制清單」走向演化與生態的整合框架。免疫反應的強弱、型別、記憶與耐受,都可視為宿主在「清除病原體」與「避免自我傷害及代謝成本」之間的權衡最佳化;病原體的毒力(virulence)也並非越強越好,而是在「複製傳播」與「不殺死宿主」之間演化出的策略。把免疫學放進演化生態學、系統生物學與數學建模的脈絡裡,最終你會看見一個比「攻防戰」更深的圖景:免疫不是一套靜態的防禦工事,而是一個在多重時間尺度上持續最佳化、永遠處於動態平衡的演化系統。理解這一點,是從「會背免疫學」邁向「會用免疫學思考」的關鍵一步。