
如果力不是只沿著肌腱傳遞,那它還能往哪裡走?
超越「肌肉拉肌腱拉骨頭」的線性模型,從橫向力傳遞、筋膜張力網、張拉整體到纖維型別與肌腱彈簧,重新理解身體如何分散與儲存力量。
如果力不是只沿著肌腱傳遞,那它還能往哪裡走?
入門篇把肌肉骨骼系統描述成一台優雅的槓桿機械:肌肉收縮、肌腱把力傳到骨頭、關節作為支點完成動作。這個「線性串聯」的圖像在教科書裡通行了一個多世紀,也確實能解釋很多事。但近三十年的解剖與生物力學研究揭露了一個更複雜、也更有趣的事實——一條肌肉所產生的力,並不會全部沿著它自己的肌腱傳出去。
研究者在動物與人體標本上做過一個關鍵實驗:把單一肌肉的近端肌腱拉緊、測量它遠端肌腱收到的力,結果發現兩端的張力對不起來。消失的那一部分力跑去哪了?答案是:它橫向穿過了包覆肌肉的結締組織,傳給了旁邊的肌肉與更遠的結構。這個現象稱為肌外力傳遞(epimuscular myofascial force transmission)。它意味著,把每條肌肉當成獨立元件、各自接到各自骨頭的傳統模型,在量化上是不完整的。本篇要帶你超越「肌肉拉肌腱拉骨頭」這條直線,進入一個由膠原蛋白網路、肌纖維代謝差異與整體張力結構共同編織的系統。
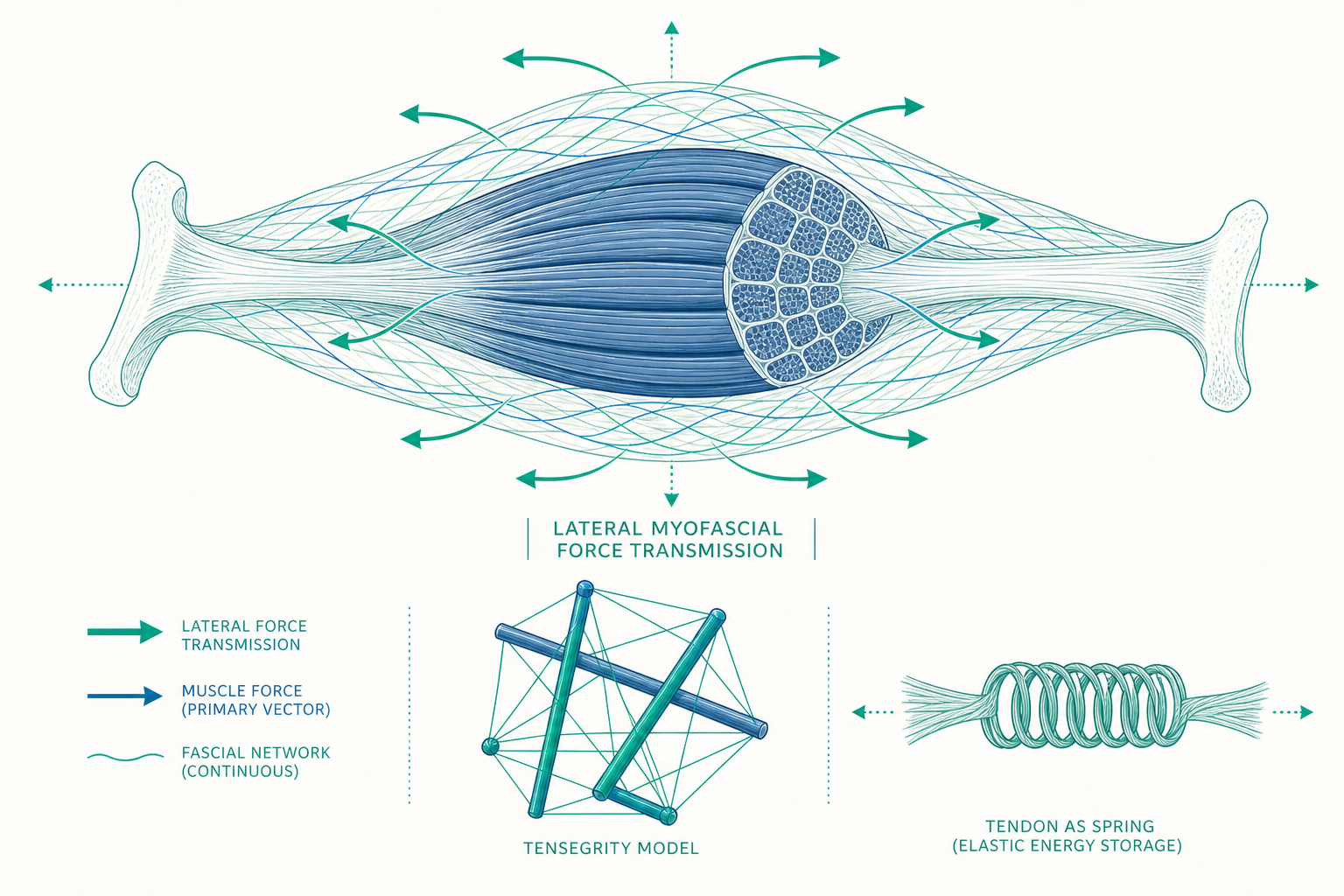
筋膜:被忽略了一個世紀的力學連續體
在傳統解剖教學裡,筋膜(fascia)常被當成解剖時要清掉、好讓「真正的構造」露出來的包裝紙。但從力學角度看,這層由膠原蛋白(collagen)與彈性蛋白(elastin)構成的結締組織,本身就是一個遍佈全身、連續不斷的張力網路。
肌肉的結締組織分為三層,由外而內層層包裹:包住整條肌肉的肌外膜(epimysium)、包住肌束的肌束膜(perimysium)、以及包住每一條肌纖維的肌內膜(endomysium)。重點在於,這三層膜並非彼此孤立——它們在分子層級互相連續,並向外與肌腱、骨膜、相鄰肌肉的筋膜接續。換句話說,一條肌纖維產生的張力,可以透過肌內膜「橫向」傳給隔壁的纖維,再經肌束膜、肌外膜往更遠的地方擴散。
這解釋了一個臨床上長期令人困惑的現象。早年的研究曾估計,肌肉與其肌腱之間的細胞連接,似乎不足以承受全力收縮時的張力——若力只走「肌纖維末端 → 肌腱」這條縱向路徑,連接點理論上應該頻繁撕裂。橫向力傳遞補上了這塊拼圖:力其實被分散到大量的側向連結上,整個肌肉像一束被網子兜住的纖維,而不是一捆各自獨立、只在末端打結的繩子。
看一個例子:為什麼按摩小腿能改善看似無關的緊繃
筋膜的連續性,在身體上構成了一些可被追蹤的「力學鏈」。其中最常被討論的是沿著身體背側、從腳底經小腿、大腿後側、脊椎兩側一路延伸到頭頂與眉骨的後表線(superficial back line)這類肌筋膜連續結構。
設想一個情境:一位學生長時間久坐,主訴下背僵緊。傳統思路會直接在下背找問題。但若把筋膜視為連續體,小腿後側與大腿後側的縮短與沾黏,也可能透過這條背側張力鏈,把張力一路「拉」到下背。臨床上,針對小腿或腳底筋膜進行放鬆,有時能改善看似無關部位的活動度——這正是把身體當成一張互相牽引的網、而非一堆獨立零件時,才容易理解的現象。
要謹慎的是,肌筋膜經線(myofascial meridians)的具體路徑與「治療甲處可解乙處」的因果強度,在實證上仍有爭議,不同研究的結論並不一致。本文介紹它,是要你掌握「力會橫向擴散、身體是連續體」這個經得起力學檢驗的核心概念,而非把任何單一經線圖當成定論。
張拉整體:身體是一座漂浮的張力結構嗎?
如果力會橫向擴散、筋膜是連續的張力網,那麼一個自然的問題是:人體的結構穩定,究竟主要靠什麼?
傳統「連續壓縮(continuous compression)」的觀點認為,身體像疊磚牆或堆積木:骨頭一塊壓著一塊,重量由上往下逐節傳遞,肌肉只是讓這堆磚動起來的馬達。但這個模型有個破綻——如果真是骨頭直接互相壓著,關節面承受的壓力會大到不合理,而且只要稍微傾斜整座「磚牆」就該垮。
另一個觀點借用了建築與雕塑裡的張拉整體(tensegrity,tensional integrity 的縮寫)概念。在張拉整體結構中,剛性的桿件(在人體即骨頭)彼此不直接接觸,而是漂浮在一張連續的張力網(在人體即肌肉、肌腱、韌帶與筋膜)之中。整個結構靠「連續的張力 + 不連續的壓縮」維持形狀:張力構件無所不在地把桿件拉住、定位,壓縮構件則被張力網懸吊著、彼此分開。
把這個概念套到人體上,骨頭就不是「互相堆疊承重」,而是被肌筋膜張力網懸吊、撐開、定位的元件。這帶來幾個有解釋力的推論:第一,負荷不會集中在單一關節面,而是被整張網分散,降低局部壓力;第二,局部的張力改變會牽動全局——拉動結構的一處,整體都會微調以重新平衡;第三,結構在受力下能彈性變形又回彈,而不是脆性地斷裂。
需要誠實指出,把人體完全化約為純粹的張拉整體結構是過度簡化的——人體骨頭在某些關節確實有壓力性的接觸,承重時關節面也確實受壓。張拉整體更像一個有用的思考框架與設計隱喻,提醒我們張力網與壓縮件如何協同分散負荷,而非一條可以精確量化全身的物理定律。它的價值在於糾正「骨頭單純堆疊承重」的直覺,而不是取代既有的關節力學。
不是所有肌纖維都一樣:纖維型別與代謝引擎
入門篇談了肌肉收縮的滑動絲機制,但有一個對運動表現與健康都至關重要的維度沒展開:肌纖維其實分成不同型別,各自配備不同的能量系統。
依收縮速度與代謝特性,骨骼肌纖維大致分為兩大類:
- 第一型(Type I,慢縮、slow-twitch)纖維:收縮慢、力量較小,但極耐疲勞。它富含粒線體(mitochondria)、肌紅蛋白(myoglobin,讓它呈現較紅的顏色)與微血管,主要靠有氧代謝(aerobic metabolism)把脂肪與葡萄糖徹底氧化產生 ATP。它是維持姿勢、長時間活動(如馬拉松、站立)的主力。
- 第二型(Type II,快縮、fast-twitch)纖維:收縮快、爆發力強,但容易疲勞。其中又可細分,譬如偏向無氧、糖解代謝(glycolytic metabolism)的亞型,靠快速分解肝醣(glycogen)取得能量,適合短跑、舉重等爆發動作;另有兼具速度與一定耐力的中間型。
人體幾乎每塊肌肉都是兩型纖維的混合,比例因人、因部位而異。維持站姿的深層肌肉慢縮比例高,而需要瞬間爆發的肌肉則快縮居多。值得釐清一個常見迷思:纖維型別的「先天比例」有相當程度由基因決定,訓練很難把一個人從以慢縮為主完全改造成以快縮為主;但訓練確實能改變纖維的代謝特性與橫斷面積——耐力訓練讓纖維變得更有氧、更抗疲勞,阻力訓練則讓快縮纖維變粗、輸出更大力量。
理解纖維型別,也就理解了能量系統的分工。短於約十秒的全力爆發,主要靠肌肉內既存的 ATP 與磷酸肌酸(creatine phosphate)系統;數秒到約兩分鐘的高強度活動,主要靠無氧糖解(會累積乳酸與氫離子,造成那種灼熱的疲勞感);超過數分鐘的持續活動,則必須仰賴有氧系統。這三套系統並非依序開關,而是始終同時運作、只是隨強度與時間改變主導者。
動手試試:用一個簡單動作分辨你的兩套引擎
找一面牆做靜態深蹲(背貼牆、大腿與地面平行)。前一、兩分鐘,你大概還撐得住,這時主要是慢縮纖維與有氧系統在工作。但隨著時間拉長,你會感到大腿前側出現明顯的灼熱與顫抖——這是快縮纖維被更大量招募、無氧糖解產物累積、運動單位開始輪替放電的綜合結果。
接著休息,改做一次盡全力的垂直跳。這一下幾乎完全是磷酸肌酸系統與快縮纖維的舞台:爆發、瞬間、然後迅速「沒電」。同一雙腿、同一批肌肉,在兩種任務下動用了截然不同的纖維與代謝路徑。你親身感受到的,正是骨骼肌作為「多引擎混合動力系統」的本質。
肌腱不只是繩子:彈性儲能與能量回收
入門篇把肌腱比喻成傳遞力量的繩索。進階一點看,肌腱與韌帶其實是會儲存與釋放能量的生物彈簧,這對運動效率有決定性影響。
當你跑步、跳躍時,落地的瞬間肌腱(尤其是阿基里斯腱)會被拉長、像彈簧一樣儲存彈性位能,接著在蹬地時把這份能量釋放出來,幫助你彈起。這種「肌肉作為馬達、肌腱作為彈簧」的協作,讓人類的行走與奔跑遠比「純靠肌肉每一步重新出力」要省能。研究估計,在跑步中,下肢肌腱的彈性回收可省下相當可觀比例的代謝能量——這是為什麼具備良好彈性的肌腱對耐力表現如此重要。
從材料力學看,肌腱表現出黏彈性(viscoelasticity):它的力學反應同時取決於變形的幅度與速率。快速施力時它較硬、能有效回彈儲能;緩慢施力時它會逐漸延展(潛變,creep)。這也解釋了臨床上一些現象,例如肌腱在反覆高速負荷下的損傷機制,與在長時間靜態牽拉下的反應並不相同。理解肌腱是速率敏感的黏彈性彈簧,而非一條死板的繩子,是進入運動傷害與復健研究的門檻概念。
重點回顧
- 力不只縱向沿肌腱傳遞:透過肌外膜、肌束膜、肌內膜的連續結構,肌肉的張力會橫向擴散到相鄰結構,這就是肌外力傳遞(epimuscular myofascial force transmission)。
- 筋膜是遍佈全身的連續張力網,把身體連成可被追蹤的力學鏈;但具體的肌筋膜經線路徑與「治甲處解乙處」的因果強度,實證上仍有爭議。
- 張拉整體(tensegrity)是有用的思考框架:骨頭像被張力網懸吊、分散負荷的壓縮件;但人體不能完全化約為純張拉整體,部分關節仍有真實的壓力接觸。
- 肌纖維分慢縮(Type I,有氧、耐疲勞)與快縮(Type II,爆發、易疲勞);先天比例多由基因決定,但訓練可改變其代謝特性與粗細。三套能量系統(磷酸肌酸/無氧糖解/有氧)始終並行、隨強度切換主導。
- 肌腱是黏彈性的生物彈簧,能儲存並回收彈性位能,其力學反應對施力速率敏感——這是運動效率與傷害機制的關鍵。
(本文為衛教與知識性質的機制說明,非個人醫療建議。任何疼痛、外傷或訓練計畫,請依個別狀況諮詢醫師、物理治療師或合格運動專業人員。)
深入探討(研究所視角)
把橫向力傳遞、筋膜連續性、纖維代謝與肌腱彈性串起來,會通往幾個目前仍活躍的研究前沿。
從肌節到全身的多尺度力學建模(multiscale modeling)。 入門篇提到的肌肉骨骼模型(如 OpenSim)多半把每條肌肉視為獨立的力源,沿著定義好的路徑收縮——這個假設在納入肌外力傳遞後就需要修正。前沿的有限元素(finite element)肌肉模型試圖把肌內膜、肌束膜的橫向耦合納入運算,讓力得以在相鄰肌肉間「滲透」。挑戰在於跨越尺度:從奈米級的橫橋、微米級的肌節、毫米級的纖維束,一路連到公分級的整塊肌肉與全身運動,每一層的力學參數都需要實驗校準,而組織的個體差異極大。這也使「肌肉冗餘問題」更複雜——當力可以橫向分流,神經系統的最佳化準則就不只是分配各肌肉的出力,還涉及整張筋膜網的張力分布。
力學生物學與細胞外基質重塑(ECM remodeling)。 橫向力傳遞之所以可能,靠的是肌纖維膜上的肌縮蛋白聚合體(dystrophin-glycoprotein complex)等結構把細胞骨架錨定到細胞外基質(extracellular matrix, ECM)。這條力學鏈一旦出問題,就直接連到病理:肌肉失養症(muscular dystrophy)正是這類錨定蛋白缺陷導致纖維膜在反覆收縮下受損。另一方面,ECM 並非被動支架,纖維母細胞(fibroblast)會感知張力、調控膠原蛋白的合成與排列,這使得制動、過度負荷、老化都會改變組織的力學性質——也讓「如何透過可控負荷引導 ECM 朝健康方向重塑」成為復健科學的核心問題。
纖維型別的可塑性與代謝健康。 慢縮/快縮的比例與代謝表型,與胰島素敏感性、第二型糖尿病風險等代謝健康指標有關聯。研究者關心的是:運動如何透過 PGC-1α 等轉錄共活化因子,驅動纖維朝更有氧、粒線體更豐富的表型轉變,以及這種「代謝重編程」能在多大程度上對抗久坐與老化帶來的代謝失調。這把骨骼肌從單純的運動器官,重新定位為調節全身代謝的關鍵組織。
跨領域連結:多模態感測與具身學習。 對教育與學習分析而言,肌肉骨骼系統的這些深層性質提供了可量測的訊號。表面肌電圖(surface EMG)能反映運動單位的招募與疲勞,慣性感測單元(IMU)能捕捉動作的協調與穩定性。當這些生理/動作模態與認知、情緒等資料整合,便有機會把「具身認知(embodied cognition)」從理論假說推進到可實證的研究——例如疲勞與姿勢如何影響專注與學習表現。這正呼應了多模態教育資料整合的方向:把身體真正納入學習的量化研究,而不是只盯著螢幕上的點擊與文字。
從一條沒有全部交給自己肌腱的張力,到一張懸吊著全身骨架的筋膜網,再到混合動力的肌纖維與會回彈的肌腱彈簧——肌肉骨骼系統的進階圖像告訴我們:身體的力學從來不是一條條獨立的直線,而是一個橫向耦合、連續分布、隨負荷重寫自己的整體網路。理解這層複雜性,正是從「會背解剖名詞」走向「能推理力如何流動」的分水嶺。