
一塊三明治去了哪裡?
從消化、吸收到代謝,追蹤食物如何被拆解成分子並轉化為身體可用的能量與材料。
一塊三明治去了哪裡?
午餐你吃下一塊夾著火腿、起司、生菜的全麥三明治。短短幾秒,它離開你的視線進入身體,接下來的幾個小時內,這份食物會被拆解成數以兆計的分子碎片,有些被運往肌肉提供能量,有些被送進肝臟改頭換面,有些則成為構築新細胞的磚塊。這一連串看似理所當然的過程,其實是一套橫跨九公尺長管道、動員數十種酵素、整合神經與內分泌訊號的精密工程。
消化系統(digestive system)的核心任務只有一句話:把「別人的分子」轉換成「你的分子」。食物中的蛋白質、脂質、碳水化合物都是大分子,無法直接穿過細胞膜,必須先被分解成小單位(胺基酸、脂肪酸、單醣),才能被吸收、運送、利用。理解這套「消化—吸收—代謝」三部曲,是看懂營養學、代謝疾病乃至許多臨床問題的基礎。
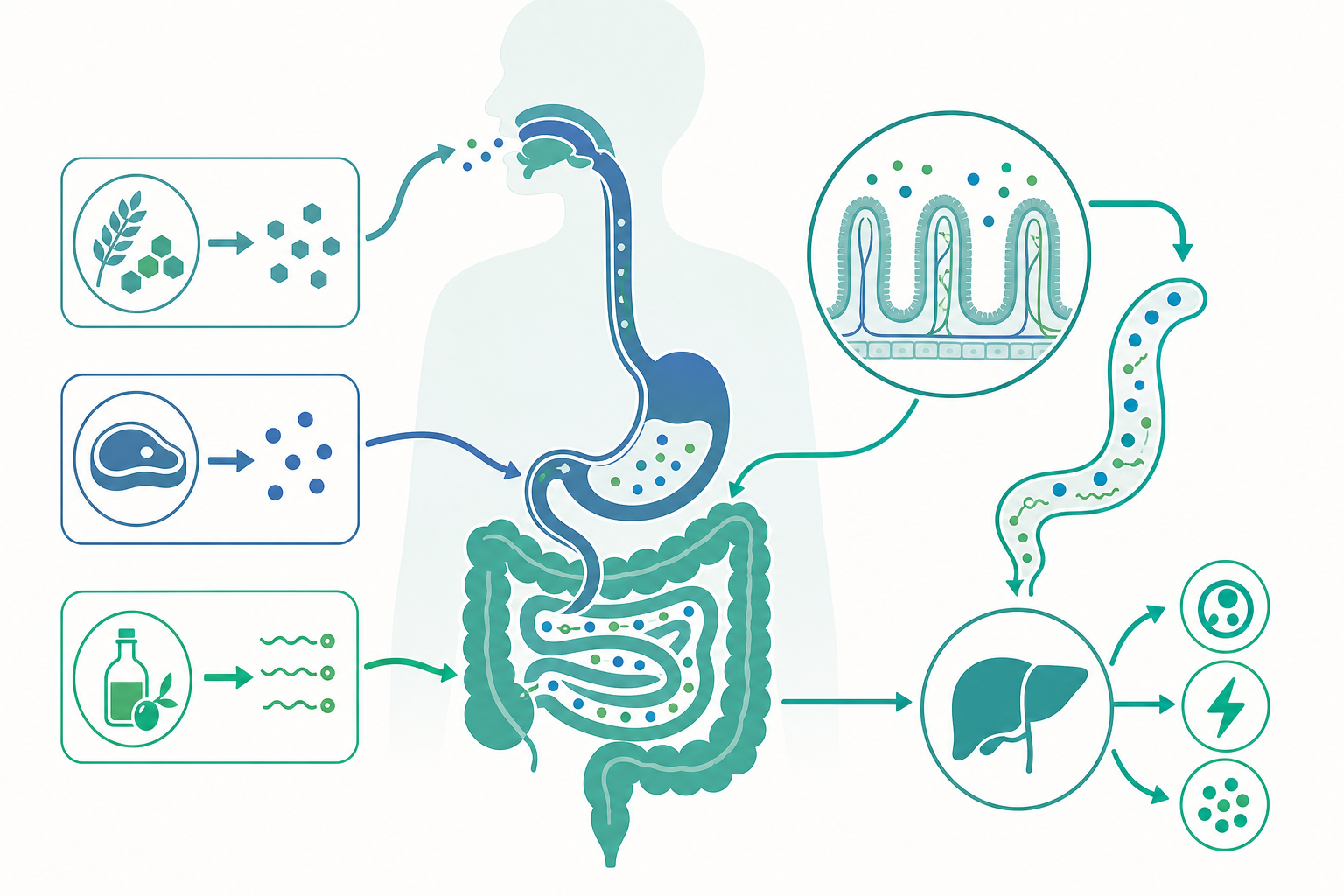
消化道:一條由內而外的「體外管道」
從解剖學角度看,消化道(gastrointestinal tract, GI tract)是一根貫穿身體的中空長管,依序由口腔、咽、食道、胃、小腸(十二指腸、空腸、迴腸)、大腸(結腸、直腸)連接到肛門。一個有趣但重要的觀念是:消化道的內腔(lumen)在拓樸學上其實是「體外」——食物在被吸收進入血液之前,從未真正進入人體內部環境。這個觀念解釋了為什麼腸道黏膜需要兼具「吸收」與「屏障」兩種角色:它要讓營養分子通過,同時阻擋細菌、毒素與未消化的大分子。
消化包含兩種方式並行運作。機械性消化(mechanical digestion)靠牙齒咀嚼、胃的攪拌、小腸的分節運動,把食物磨碎、混合並增加表面積;化學性消化(chemical digestion)則靠酵素(enzyme)把化學鍵切斷。兩者相輔相成:機械性消化把食物打散,讓酵素有更多接觸面可以作用。
消化道的運動由腸神經系統(enteric nervous system, ENS)主導,這套被稱為「第二大腦」的神經網路含有約五億個神經元,能在不依賴中樞神經的情況下協調蠕動(peristalsis)與分泌。自主神經系統則扮演調節角色:副交感神經(迷走神經為主)促進消化分泌與蠕動,交感神經則在壓力或運動時抑制消化。這也是為什麼焦慮緊張時容易出現腸胃不適。
從口腔到胃:消化的起點
消化從你看到、聞到食物的那一刻就已啟動。頭期反應(cephalic phase)透過迷走神經預先刺激唾液與胃液分泌,讓身體「準備好」接收食物。
唾液中的唾液澱粉酶(salivary amylase, ptyalin)開始分解澱粉,把長鏈多醣切成較短的麥芽糖與糊精。不過唾液澱粉酶在酸性的胃中很快失活,因此口腔內的澱粉消化只是序曲。
食物經吞嚥、由食道蠕動送入胃。胃是一個強酸性的攪拌槽,胃壁的壁細胞(parietal cells)分泌鹽酸(HCl),把胃內 pH 維持在約 1.5–3.5。如此強酸有三個功能:使蛋白質變性(denature)以利酵素作用、活化消化酵素、殺滅多數隨食物進入的微生物。主細胞(chief cells)分泌的胃蛋白酶原(pepsinogen)在酸性環境下被活化為胃蛋白酶(pepsin),開始將蛋白質切成較小的多肽片段。值得注意的是,碳水化合物與脂質在胃中幾乎沒有被化學消化,胃對它們主要做機械性攪拌,形成半流質的食糜(chyme)。
胃壁本身為什麼不會被自己的強酸與蛋白酶消化?關鍵在於黏液細胞分泌的黏液—碳酸氫鹽屏障(mucus-bicarbonate barrier),它在胃壁表面形成一層中性緩衝膜。當這層保護被破壞(例如幽門螺旋桿菌感染或長期使用非類固醇消炎藥 NSAIDs 抑制保護性前列腺素),就可能形成消化性潰瘍(peptic ulcer)。
小腸:消化與吸收的主戰場
食糜分批通過幽門進入十二指腸,這裡才是化學消化真正的高潮,也是絕大多數營養吸收發生的地方。三股力量在此匯合。
胰臟(pancreas)的外分泌提供一整套消化酵素:胰澱粉酶(pancreatic amylase)繼續分解碳水化合物;胰脂酶(pancreatic lipase)負責脂質;蛋白酶(如胰蛋白酶 trypsin、胰凝乳蛋白酶 chymotrypsin)處理蛋白質。胰臟同時分泌大量碳酸氫鹽(bicarbonate),中和來自胃的酸性食糜,把腸腔 pH 拉回中性偏鹼,這是胰酵素發揮活性的必要條件。
肝臟(liver)製造、膽囊(gallbladder)儲存的膽汁(bile)雖不含酵素,卻對脂質消化至關重要。膽汁中的膽鹽(bile salts)是兩親性分子,能把大的脂肪滴乳化(emulsification)成微小脂肪滴,大幅增加脂酶可作用的表面積。沒有膽汁,脂肪消化效率會嚴重下降。
小腸黏膜本身也參與最後一步消化。腸壁上皮細胞的刷狀緣(brush border)含有刷狀緣酵素,例如分解雙醣的乳糖酶(lactase)、蔗糖酶(sucrase)、麥芽糖酶(maltase),以及把小肽切成胺基酸的肽酶(peptidase),完成消化的「收尾」。乳糖不耐(lactose intolerance)正是因為乳糖酶活性不足,未被分解的乳糖進入大腸後被細菌發酵,產生氣體與滲透性腹瀉。
小腸的吸收效率源自驚人的表面積設計:環狀皺襞(plicae circulares)、絨毛(villi)與微絨毛(microvilli)三層放大結構,使小腸內表面積達到約 200–250 平方公尺,相當於一個網球場。營養分子在此被吸收進入絨毛內的微血管或乳糜管(lacteal)。
三大營養素的吸收路徑
不同營養素被吸收進入體內的「門」並不相同,理解這點有助於看懂代謝與運送邏輯。
碳水化合物最終以單醣(葡萄糖 glucose、半乳糖 galactose、果糖 fructose)形式被吸收。葡萄糖與半乳糖透過鈉—葡萄糖共同運輸蛋白(SGLT1)進行次級主動運輸,借助鈉離子梯度逆濃度吸收;果糖則靠 GLUT5 被動擴散。這些單醣進入微血管,匯入肝門靜脈(hepatic portal vein)直送肝臟。
蛋白質被分解為胺基酸與小肽後,同樣經由鈉依賴或氫依賴的轉運蛋白被吸收,進入肝門靜脈系統。
脂質的路徑最為特別。脂肪酸與單酸甘油酯先與膽鹽形成微膠粒(micelle),運送到刷狀緣後被吸收進細胞,在細胞內重新組裝成三酸甘油酯,並包裹成乳糜微粒(chylomicron)。由於乳糜微粒體積太大無法進入微血管,它們改走淋巴系統:先進入絨毛的乳糜管,經淋巴管最終由胸管匯入鎖骨下靜脈進入血液循環。這也是為什麼脂溶性物質(包括脂溶性維生素 A、D、E、K)的吸收依賴正常的脂肪消化。
大腸與腸道菌群:常被低估的末段
當食糜抵達大腸時,大部分營養與水分其實已被小腸吸收。大腸的主要工作是回收水分與電解質,把剩餘殘渣濃縮成糞便。若大腸蠕動過快、水分來不及回收就是腹瀉;蠕動過慢、水分回收過度則造成便秘。
但大腸絕非單純的「廢物處理場」。棲息其中的腸道菌群(gut microbiota)多達數百種、數量以兆計,它們發酵小腸無法消化的膳食纖維,產生短鏈脂肪酸(short-chain fatty acids, SCFAs,如丁酸 butyrate),這些是結腸上皮細胞的重要能量來源,並具有調節免疫與發炎的作用。腸道菌也合成部分維生素 K 與 B 群。近年研究顯示,腸道菌群的組成與肥胖、第二型糖尿病、免疫疾病甚至情緒(透過腸—腦軸 gut-brain axis)都有關聯,是當代生醫研究的熱門前沿。
代謝:吸收之後的分子重組
吸收只是把營養分子搬進體內,真正決定它們命運的是代謝(metabolism)——細胞層級的化學轉換網路。代謝分為兩大方向:分解代謝(catabolism)把大分子拆解並釋放能量(以 ATP 形式儲存),合成代謝(anabolism)則消耗能量去建構新分子。
肝臟是代謝的中央調度站。經肝門靜脈而來的營養素首先在此「過關」:葡萄糖過多時被合成為肝醣(glycogen)儲存,或轉化為脂肪;血糖不足時則分解肝醣或進行糖質新生(gluconeogenesis)釋放葡萄糖。胺基酸在肝臟被用於合成蛋白質,多餘的部分經去胺作用(deamination)移除含氮基團,產生的氨經尿素循環(urea cycle)轉為尿素由腎臟排出。
血糖的恆定由胰島素(insulin)與升糖素(glucagon)這對拮抗激素調控:餐後血糖升高,胰島素促進細胞攝取葡萄糖並儲存;空腹時升糖素則動員肝醣與糖質新生維持血糖。當這套調控失靈——胰島素分泌不足或細胞對胰島素反應變差(胰島素阻抗 insulin resistance)——就導向糖尿病(diabetes mellitus)。這正是消化吸收與代謝環環相扣的臨床縮影:問題未必出在「吃」,而可能出在吸收後分子如何被處理。
看一個例子
一位 35 歲女性近半年常在喝牛奶或吃冰淇淋後約 30 分鐘到 2 小時內出現腹脹、腸鳴與水樣腹瀉,但吃優格或起司時症狀輕微。她沒有發燒、體重穩定、糞便無血。
從消化生理推理:乳製品的主要醣類是乳糖(lactose),屬於雙醣,必須靠小腸刷狀緣的乳糖酶水解為葡萄糖與半乳糖才能被吸收。若乳糖酶活性不足,未被分解的乳糖滯留在腸腔,一方面造成滲透性效應把水分拉進腸道,另一方面進入大腸後被菌群發酵,產生氫氣、二氧化碳與短鏈脂肪酸,於是出現腹脹、排氣與腹瀉。為什麼優格與硬起司症狀較輕?因為發酵過程與熟成已讓部分乳糖被細菌代謝掉,殘餘乳糖較少。
臨床上可用氫氣呼氣試驗(hydrogen breath test)輔助判斷:攝取乳糖後若呼氣中氫氣明顯上升,支持乳糖在大腸被發酵的推論。處置原則是調整乳糖攝取量、選擇低乳糖或無乳糖製品,或補充外源性乳糖酶,而非完全排斥所有鈣質來源。
這個案例示範了如何用「消化—吸收—代謝」的邏輯,把一個日常症狀拆解到分子層級。它也提醒:成人乳糖酶活性下降是全球多數族群的常態生理變化,並非疾病。(本文為教育用途,不構成個人醫療建議;若有持續腸胃症狀,請諮詢合格醫療人員。)
重點回顧
- 消化系統的核心任務是把食物中的大分子(蛋白質、脂質、碳水化合物)分解成可吸收的小單位,再透過吸收與代謝轉化為身體可用的能量與材料。
- 化學消化分階段進行:口腔(澱粉)、胃(蛋白質、強酸環境)、小腸(三大營養素的主戰場,整合胰酵素、膽汁與刷狀緣酵素)。
- 小腸靠皺襞、絨毛、微絨毛三層結構放大表面積至約 200 平方公尺,是吸收的主要場所;脂質走淋巴系統,醣類與胺基酸走肝門靜脈。
- 大腸負責回收水分與電解質,腸道菌群發酵纖維產生短鏈脂肪酸,與免疫、代謝乃至腸—腦軸密切相關。
- 吸收之後的代謝由肝臟主導,並由胰島素與升糖素調控血糖恆定;此調控失衡即連結到糖尿病等代謝疾病。
深入探討(研究所視角)
對於進一步鑽研的學習者,消化與代謝的交界處正是當代生醫研究最活躍的地帶。
腸—腦軸與菌群代謝體(microbiome metabolomics):腸道菌群不只是消化的配角,它們透過短鏈脂肪酸、神經傳導物質前驅物(如色胺酸代謝物)與迷走神經傳訊,雙向影響中樞神經與情緒。研究方法上,16S rRNA 定序與鳥槍法總體基因體學(shotgun metagenomics)讓我們得以描繪菌群組成,而代謝體學(metabolomics)則進一步量化菌群產物。一個關鍵的研究挑戰是區分「相關」與「因果」——糞菌移植(fecal microbiota transplantation)的動物與人體試驗正試圖回答這個問題。
腸道荷爾蒙與食慾/血糖調控:腸道是體內最大的內分泌器官。腸促胰島素(incretins,如 GLP-1、GIP)在進食後由腸道細胞分泌,刺激胰島素釋放並抑制食慾。近年大放異彩的 GLP-1 受體促效劑(如 semaglutide)原本用於治療第二型糖尿病,後來在體重管理上展現顯著效果,正是把「消化道訊號」轉化為「代謝治療」的典範。這提醒我們:消化系統與內分泌、代謝的界線其實是人為劃分的。
營養素吸收的個體差異與精準營養(precision nutrition):同樣一份餐點,不同人的餐後血糖反應可以差異極大。以色列 Weizmann 研究團隊(Zeevi 等人,2015,Cell)以連續血糖監測結合機器學習,發現個人化的血糖反應可由菌群、生活型態與飲食特徵預測,挑戰了「升糖指數人人適用」的傳統觀念,開啟精準營養的研究方向。
方法學提醒:營養與代謝研究常受限於觀察性設計的干擾因子(confounding)與飲食回報的測量誤差。閱讀文獻時,宜留意研究是隨機對照試驗還是觀察性世代、樣本量與族群代表性、以及機制證據(mechanistic evidence)是否支持流行病學關聯。把分子機制與群體實證並置檢驗,才是嚴謹的營養科學思維。