
為什麼荷爾蒙不是「水龍頭」,而是「鼓點」?
荷爾蒙傳遞的不只是「多少」,更是「什麼時候、以什麼節奏」——從脈衝頻率編碼、HPA 壓力軸的雙受體剎車,到把內分泌當成會振盪的動態系統來理解。
為什麼荷爾蒙不是「水龍頭」,而是「鼓點」?
如果你讀過入門篇,你已經知道內分泌系統用荷爾蒙在血液裡傳遞化學訊息,也知道負回饋(negative feedback)像恆溫器一樣維持恆定。但這裡有一個入門課本很少直說、卻足以顛覆直覺的事實:大多數荷爾蒙並不是被「持續穩定地」分泌的,而是一陣一陣、像鼓點般脈衝式(pulsatile)地釋放。
考慮一個臨床上真實發生過的怪事:醫師想用性釋素(GnRH,gonadotropin-releasing hormone)治療某些不孕症患者,於是給予「持續、穩定」的 GnRH——濃度拉得高高的、整天不間斷。結果不但沒有刺激性腺,反而把整條軸關掉了。同一個分子,脈衝給藥能催動生殖,持續給藥卻變成化學去勢(這正是攝護腺癌療法 GnRH agonist 的作用原理)。
訊號的「時間結構」竟然比「平均濃度」更關鍵。這就是進階內分泌學的入口:荷爾蒙傳遞的不只是「多少」,更是「什麼時候、以什麼節奏」。要理解這件事,我們得把內分泌系統當成一個有節律、有延遲、有非線性的動態系統(dynamical system)來看。
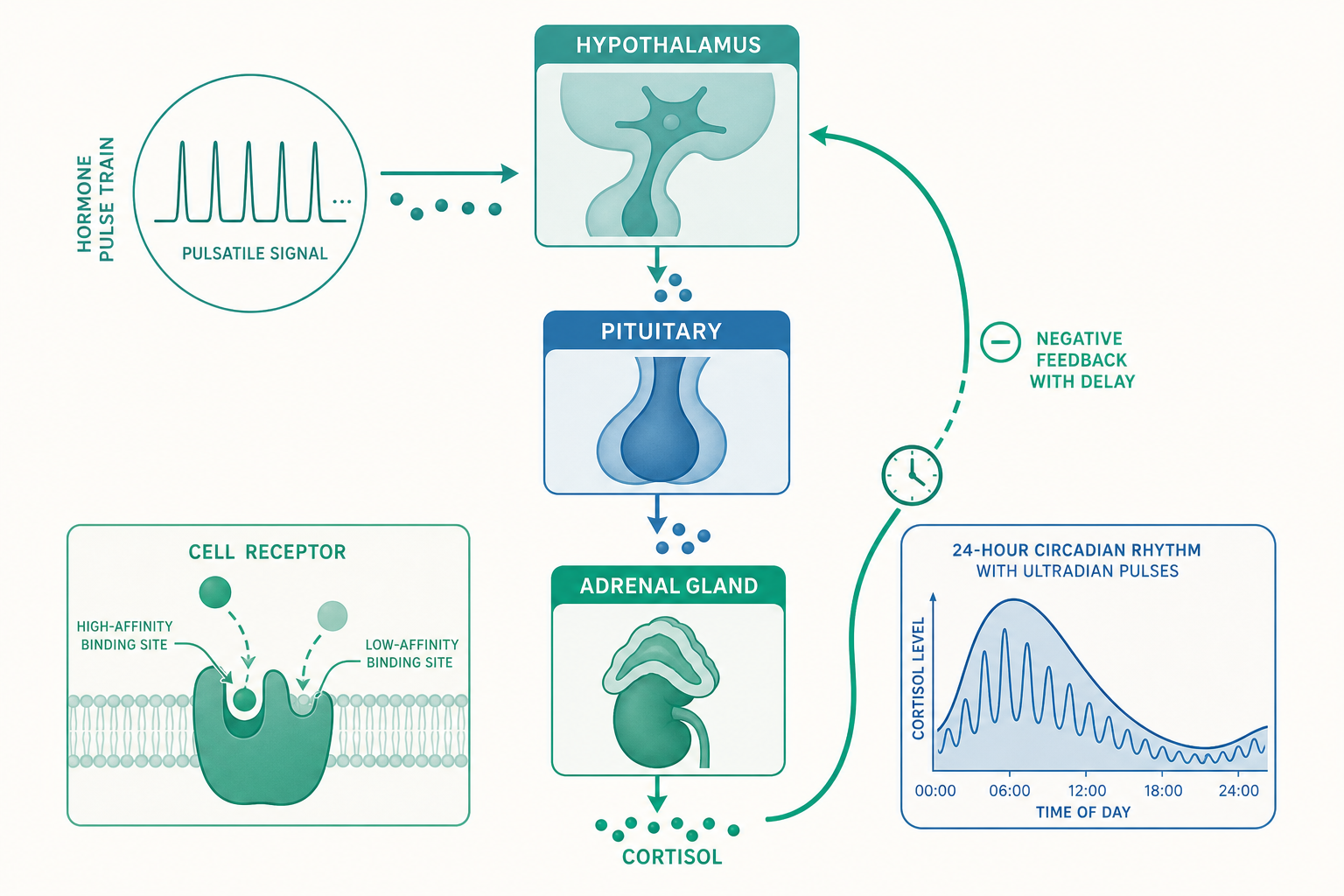
脈衝、節律與頻率編碼
入門篇提過皮質醇(cortisol)有日節律,清晨高、深夜低。但這只是表層。如果你用高時間解析度的採血(每 10 分鐘抽一次)去追蹤,會看到日節律這條「大波浪」之上,還疊著每小時數次的超日節律(ultradian rhythm)小脈衝。荷爾蒙濃度的真實曲線不是平滑的正弦波,而是鋸齒狀的一串脈衝,再被一個 24 小時的緩慢起伏調制。
為什麼身體要這麼費事地「打鼓」,而不乾脆穩定供應?關鍵在於受體的去敏感化(desensitization)與下調(down-regulation)。細胞表面的受體若被荷爾蒙長時間佔據,會被內吞、磷酸化失活、甚至減少數量——這是細胞保護自己不被訊號淹沒的本能。脈衝式分泌讓受體在脈衝之間有時間「重設」,恢復敏感度。於是脈衝的頻率(frequency)就成了一個獨立的資訊維度:細胞可以「數脈衝有多密」來判讀指令,而不只是「測濃度有多高」。這叫頻率編碼(frequency coding)。
GnRH 是最乾淨的例子。下視丘以大約每 60–90 分鐘一次的頻率脈衝釋放 GnRH,腦下垂體據此分泌兩種促性腺激素:
- 快頻率脈衝偏向促進黃體生成素(LH,luteinizing hormone)。
- 慢頻率脈衝偏向促進濾泡刺激素(FSH,follicle-stimulating hormone)。
整個月經週期的荷爾蒙交響樂,本質上就是 GnRH 脈衝頻率被動態調整的結果。所以前面那個「持續給藥反而關機」的謎題就解開了:持續、無脈衝的 GnRH 等於把頻率資訊抹平成一條直線,腦下垂體的受體被持續佔據而徹底去敏感,整條軸隨之沉默。
這帶出一條進階主線:內分泌訊號是時間序列,不是單一數值。 同樣一張抽血報告,少了時間結構,可能完全誤判。
壓力的內分泌學:HPA 軸與它的雙速度回饋
入門篇用甲狀腺軸(HPT)示範負回饋,但壓力反應所走的下視丘—腦下垂體—腎上腺軸(HPA axis,hypothalamic-pituitary-adrenal axis),是一條設計得更精巧、也更值得深究的軸。它是我們理解「慢性壓力如何傷身」的生理骨架。
HPA 軸的串接是這樣:
- 下視丘感知到壓力(生理或心理),分泌促腎上腺皮質素釋素(CRH,corticotropin-releasing hormone)。
- CRH 刺激腦下垂體分泌腎上腺皮質刺激素(ACTH,adrenocorticotropic hormone)。
- ACTH 經血液抵達腎上腺皮質,刺激分泌皮質醇。
- 皮質醇升高後,反向抑制下視丘的 CRH 與腦下垂體的 ACTH——典型負回饋。
但精妙之處在於這個負回饋有兩種速度,由兩種不同的受體分工:
- 礦皮質素受體(MR,mineralocorticoid receptor):對皮質醇親和力高,在皮質醇基礎濃度下就大半被佔據。它負責維持日常的基礎調節與晝夜節律的「設定點」。
- 糖皮質素受體(GR,glucocorticoid receptor):對皮質醇親和力低,平時多半空著,只有在壓力造成皮質醇大量飆升時才大量被佔據,啟動強力的負回饋煞車,把反應拉回來。
這個「高親和力管日常、低親和力管緊急」的雙受體設計,讓 HPA 軸同時具備靈敏的基礎調控與強力的緊急剎車。理解 MR/GR 的分工,是理解為什麼慢性壓力下系統會「校準失靈」的關鍵——當皮質醇長期偏高,GR 持續被佔據而下調,負回饋剎車逐漸失靈,皮質醇就更難被拉回來,形成惡性循環。
看一個例子:用回饋邏輯讀懂「庫欣 vs 愛迪生」
入門篇教過用「T₄ 與 TSH 的高低組合」定位甲狀腺病灶。同樣的負回饋推理,可以拆解兩個經典的腎上腺疾病。
病人 A:皮質醇高、ACTH 也高。病人 B:皮質醇高、ACTH 卻低。問題各出在哪一層?
依 HPA 軸負回饋邏輯:皮質醇高時,ACTH 理應被抑制而降低。
- 病人 A(皮質醇高、ACTH 高):負回饋「失效」了——上游還在拚命分泌 ACTH。這指向上游驅動,例如腦下垂體長了會分泌 ACTH 的腫瘤(庫欣氏病,Cushing's disease)。
- 病人 B(皮質醇高、ACTH 低):負回饋其實正常運作——皮質醇高,所以 ACTH 被壓低了。問題出在下游自己亂跑,例如腎上腺腫瘤自主分泌皮質醇(不受 ACTH 控制)。
反過來看皮質醇不足的愛迪生氏病(Addison's disease):若是腎上腺本身壞掉(原發性),皮質醇低、ACTH 代償性升高;若是腦下垂體 ACTH 不夠(次發性),則皮質醇低、ACTH 也低。
你會發現這套「看上下游兩個數字的高低組合」是內分泌學通用的定位術——對甲狀腺軸、腎上腺軸、性腺軸都成立。掌握它,你就握住了內分泌診斷的一把萬用鑰匙。
(提醒:以上為一般病理生理學說明與教學推理,並非個人醫療診斷或建議。任何症狀與檢驗的判讀,請交由你的醫療團隊。)
從「恆定」到「應變穩態」:allostasis 與累積耗損
入門篇的核心觀念是恆定(homeostasis)——把變數拉回固定設定點。但現代壓力生理學認為這個圖像太靜態了。身體真正在做的,往往不是「守住一個不變的設定點」,而是主動預測、提前調整設定點以應付即將到來的需求。這個更動態的概念叫應變穩態(allostasis)——「透過改變來達成穩定」。
例如你早晨醒來前,皮質醇就已經開始爬升,這不是對「已發生的壓力」做反應,而是身體預先動員資源,準備迎接一天的活動。設定點本身是會隨情境移動的。
但應變是有代價的。每一次動員壓力荷爾蒙、升高血壓、調動血糖,都對身體造成微小磨損。當壓力是急性、偶發的,系統用完就關,不留後遺;但當壓力變成慢性、反覆、無法關閉時,這些反應的累積成本就會堆疊起來,稱為應變負荷(allostatic load),極端時稱 allostatic overload。
應變負荷的生理表現是一連串可量測的失調:皮質醇日節律變平(清晨不夠高、深夜不夠低)、GR 負回饋遲鈍、發炎指標上升、HRV(心率變異性,heart rate variability)下降、胰島素敏感度惡化。這把看似分屬不同系統的問題——睡眠差、代謝差、情緒低落、免疫失衡——統一在「慢性壓力造成內分泌調控網路長期偏移」這條主線下。
對 Uedu 這樣整合多模態學習資料的平台,這個觀念格外有意義。穿戴裝置量到的 HRV、睡眠、壓力指數,背後正是 HPA 軸與自律神經系統的活動印記。一個學生考前長期熬夜、皮質醇節律被打亂,反映在生理數據上的,不只是「累」,而是一個可被追蹤、被理解的內分泌動態過程。
把荷爾蒙網路當成動態系統來建模
進階內分泌學最迷人的轉向,是從「敘述性的箭頭圖」走向「可計算的數學模型」。一條軸有分泌、有延遲、有非線性回饋,這正是動態系統理論擅長分析的對象。幾個值得認識的概念:
-
延遲造成振盪(delay-induced oscillation):荷爾蒙從分泌到抵達標的、到產生效果、再到回饋抑制上游,整個迴路有時間延遲。一個帶延遲的負回饋系統,數學上天生傾向振盪而非靜止——這從根本上解釋了為什麼荷爾蒙會脈衝,而不是停在一條死水般的穩定線上。脈衝不是 bug,是帶延遲負回饋系統的自然行為。
-
雙穩態與切換(bistability & switching):當回饋帶有夠強的非線性(例如某些正回饋環節),系統可能出現兩個穩定狀態,並在特定閾值下「咔嗒」一聲切換過去。月經週期中期 LH 的劇烈飆升(LH surge)——由雌激素在高濃度下短暫轉為正回饋所驅動——就帶有這種開關式切換的味道。入門篇提過正回饋必須有明確終點,這裡正是它在週期性事件中被精準運用的範例。
-
頻率解調(frequency demodulation):細胞如何把「脈衝頻率」翻譯成「反應強度」?這牽涉受體與下游訊號分子的活化/去活化動力學的時間常數。當脈衝間隔與這些時間常數匹配時,細胞反應最大——本質上細胞是一個對特定頻率「共振」的濾波器。
動手試試:自己推一條會振盪的迴路
你不需要解微分方程,也能用紙筆走一遍「為什麼負回饋會振盪」的直覺。想像一條有延遲的迴路:
- 假設此刻荷爾蒙偏低。負回饋的抑制被解除,上游於是全力分泌。
- 但效果有延遲。等荷爾蒙真正升高、訊號傳回上游時,濃度其實已經衝過頭了(因為上游剛剛全力踩了好一陣油門)。
- 濃度過高,負回饋強力踩剎車,上游完全停止分泌。
- 一樣有延遲。等濃度真正掉下來、訊號傳回上游時,又已經跌過頭——回到步驟 1。
你會發現,只要回饋存在「反應延遲」,系統就無法精準停在設定點,而是不斷地衝過頭、修正過頭,繞著設定點上下振盪。把這個直覺放大,就是真實荷爾蒙脈衝背後的數學骨架。這也說明了一件事:有時候「不穩定的振盪」才是健康的常態,過度平滑反而是病態(例如皮質醇節律變平)。
重點回顧
- 荷爾蒙多為脈衝式分泌,脈衝的頻率本身就是一個資訊維度(頻率編碼);GnRH 是最佳範例,持續給藥反而會使受體去敏感、關閉整條軸。
- HPA 壓力軸用 MR/GR 雙受體實現雙速度負回饋:高親和力的 MR 管日常基礎與節律,低親和力的 GR 管緊急剎車;慢性高皮質醇會使 GR 下調、剎車失靈。
- 「上游/下游兩個數字的高低組合」是內分泌診斷的萬用定位術,對甲狀腺、腎上腺、性腺各軸通用(如以皮質醇+ACTH 區分庫欣氏病與腎上腺腫瘤)。
- 應變穩態(allostasis)比靜態恆定更貼近現實:身體會預測性地移動設定點;慢性壓力累積成應變負荷,表現為皮質醇節律變平、GR 遲鈍、HRV 下降、代謝惡化。
- 帶延遲的負回饋天生傾向振盪,脈衝是動態系統的自然產物而非缺陷;理解荷爾蒙網路越來越仰賴動態系統與計算模型的視角。
深入探討(研究所視角)
走到研究前沿,內分泌學與系統生物學、計算神經內分泌學、精準醫療的邊界已經模糊。幾個值得追蹤的方向:
第一,神經內分泌脈衝產生器的分子基礎。 GnRH 脈衝由下視丘一群表現 kisspeptin、neurokinin B 與 dynorphin 的神經元(合稱 KNDy 神經元)構成的微迴路驅動,這個發現重寫了生殖內分泌學。臨床上,kisspeptin 類似物與 neurokinin B 受體拮抗劑已進入治療多囊性卵巢症候群(PCOS)、更年期熱潮紅的研究與應用。脈衝產生器從「黑盒子」變成可藥理操控的標的,是近十年最重要的進展之一。
第二,糖皮質素訊號的時間藥理學(chronopharmacology)。 既然皮質醇是脈衝+日節律,那麼治療腎上腺功能不足時,用「定時定量一天兩三次」的傳統給藥,其實無法重現生理脈衝。新一代的調節釋放型氫化可體松(modified-release hydrocortisone)試圖模擬生理節律。更廣地說,「在對的時間給藥」(chronotherapy)正成為一個橫跨內分泌、腫瘤、心血管的研究領域——因為受體表現與藥物代謝本身也有日節律。
第三,糖皮質素的基因組調控與組織特異性。 GR 是一個轉錄因子,被皮質醇活化後進入細胞核,可調控數百到上千個基因,且在不同組織(肝、肌肉、免疫細胞、腦)開啟的基因組完全不同。這解釋了為什麼長期使用類固醇藥物會有那麼廣泛的副作用(血糖升高、骨質流失、免疫抑制、情緒改變),也催生了選擇性糖皮質素受體調節劑(SEGRM)的研發——希望保留抗發炎效果、避開代謝副作用。表觀遺傳學(epigenetics)研究更發現,早年壓力可透過 DNA 甲基化「銘印」在 GR 基因的啟動子上,影響成年後的壓力反應,把社會經驗與分子調控連了起來。
第四,多模態整合與計算內分泌學。 把高頻採樣的荷爾蒙資料、穿戴裝置的 HRV/睡眠/活動、以及環境暴露資料放進同一個動態模型,是當前神經內分泌學與數位健康的交會點。這正呼應 Uedu 的 Educational Omics 框架:PhysioNeuromics 維度量測的生理節律,本質上就是內分泌動態系統在體表留下的可觀測投影。對有志的學生,建議三條路徑並進:(一)分子層級——精讀核受體結構與訊號傳導動力學;(二)系統層級——學習用延遲微分方程與穩定性分析描述回饋迴路的振盪、雙穩態與共振;(三)轉譯層級——追蹤從 KNDy 神經元、時間藥理到 GLP-1/GIP 雙重促效劑等代謝新藥的臨床進展。
內分泌系統最深的一課或許是:健康往往不在於把每個數值「鎖死」在一點,而在於維持一組恰到好處的節律與彈性。一個還會脈衝、還會隨晝夜起伏、還能在壓力後乾淨地關閉的系統,才是有韌性的系統。讀懂荷爾蒙的時間結構,就是讀懂身體如何在變動中保持穩定。