
空腹的腸子在做什麼?——當你沒在吃飯,消化道並沒有真的休息
從起搏慢波、腸神經系統、腸—腦軸到膽酸訊號與黏膜免疫,把消化道理解為一個有節律、半自主且向全身發訊的整合器官。
空腹的腸子在做什麼?——當你沒在吃飯,消化道並沒有真的休息
入門篇把消化道描述成一條「食物進來才開工」的加工管線:吃下三明治、酵素切割、絨毛吸收,事情就辦完了。這個圖像沒有錯,但它漏掉了一個關鍵事實——消化道是一個具有自主節律、會自己排程、且不斷向全身發送荷爾蒙訊號的器官,它的活動並不以「有沒有食物」為唯一開關。
來想一個違反直覺的問題:當你睡著、超過十二小時沒進食,胃與小腸是不是就靜止了?答案是否定的。空腹時,消化道會啟動一套被稱為移行性運動複合波(migrating motor complex, MMC)的週期性收縮,大約每 90 至 120 分鐘掃過整條腸道一次,像清道夫一樣把殘渣、脫落的上皮細胞與細菌推往下游。臨床上有些人 MMC 功能受損,未被清掃的內容物滯留,便容易出現小腸細菌過度生長(small intestinal bacterial overgrowth, SIBO)。換言之,消化道有「進食模式」與「禁食模式」兩套程式,而入門篇只講了前者。
進階篇要把消化系統重新定位為三件事的整合:一個有電生理節律的運動器官、一個擁有自己神經系統的半自主單位,以及一個用膽酸與腸道荷爾蒙對全身下指令的內分泌中樞。理解這三層,你才會明白為什麼便祕、糖尿病、減重藥物與帕金森氏症,竟然可以在「腸」這個交會點上連起來。
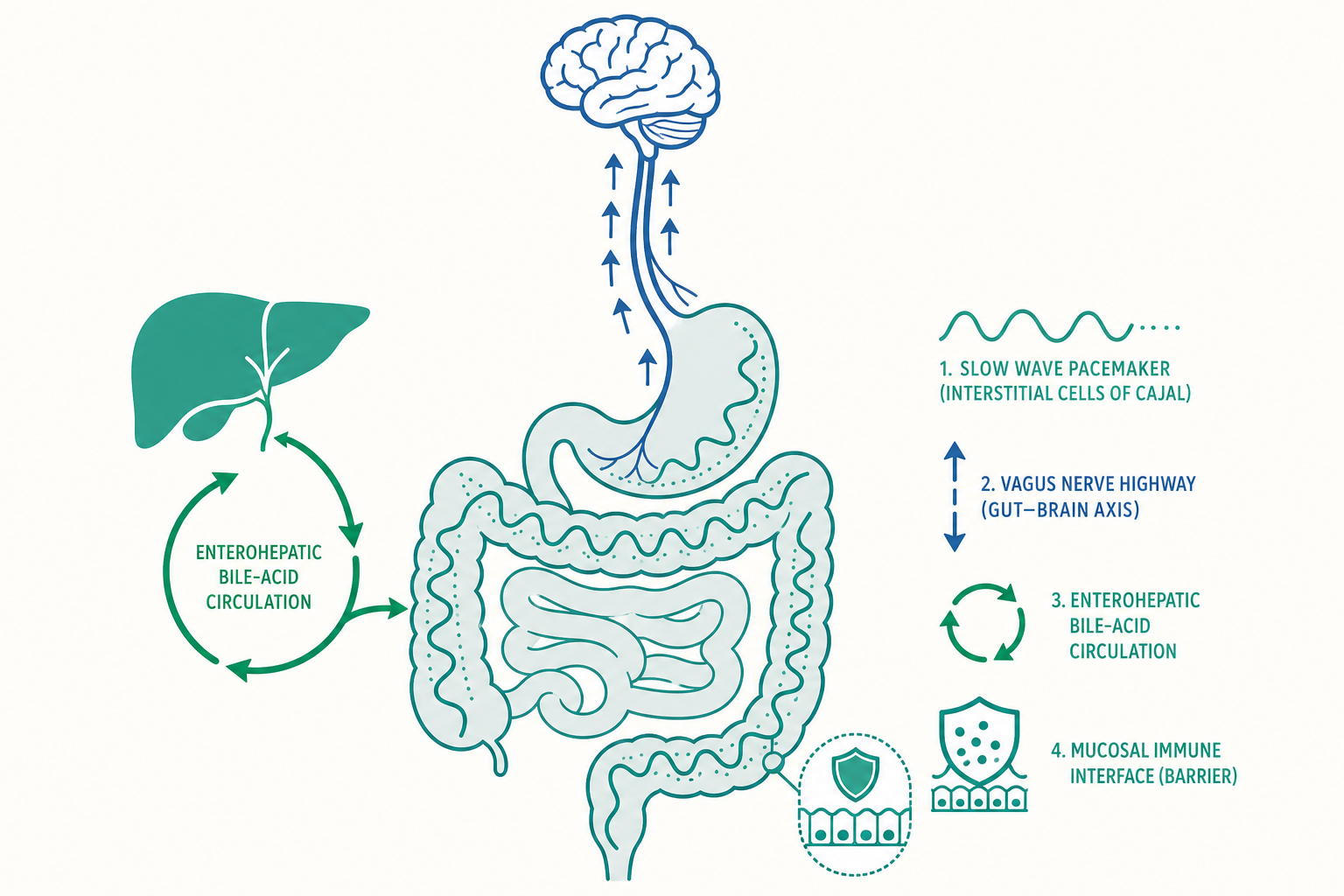
腸道的「心律」:慢波、Cajal 間質細胞與起搏節律
消化道的蠕動(peristalsis)不是隨機的肌肉抽動,而是建立在一套類似心臟的電生理起搏系統之上。
關鍵角色是Cajal 間質細胞(interstitial cells of Cajal, ICC)。這群分布在平滑肌層之間的特化細胞,扮演消化道的「起搏器」,會自發產生節律性的膜電位震盪,稱為慢波(slow waves)。慢波本身通常不直接引發收縮——它是一種「底層節拍」,把整段腸壁的興奮性調到同步。只有當慢波的去極化高峰,疊加上來自神經或荷爾蒙的興奮訊號、使膜電位越過閾值產生動作電位(spike potentials)時,平滑肌才真正收縮。
這個設計有幾個深刻的含義。第一,慢波頻率決定了該段腸道「最多能收縮多快」:胃約每分鐘 3 次、十二指腸約 12 次、迴腸約 8 次。這個由口側往肛側遞減的頻率梯度,正是食糜被淨向下游推送、而不是上下亂竄的物理基礎。第二,ICC 一旦受損(例如在某些糖尿病性胃輕癱 gastroparesis 與慢性假性腸阻塞的病人身上),慢波會變得紊亂,運動隨之失調。把腸道想成「一個有節律、需要起搏細胞、會因起搏系統故障而心律不整的器官」,比把它想成單純的肌肉管,更貼近真實。
看一個例子:胃輕癱為什麼會「吃一點點就飽、還反覆嘔吐隔餐食物」
一位罹患第一型糖尿病二十年的患者,近一年常在飯後飽脹、噁心,偶爾嘔吐出數小時前甚至前一餐的食物,胃鏡卻找不到阻塞。這是典型的糖尿病性胃輕癱。
從進階機制推理:長期高血糖會損害支配胃的迷走神經(自主神經病變),同時破壞胃竇的 ICC 起搏網路與抑制性的一氧化氮(NO)神經元。結果是胃竇收縮無力、幽門協調失常,胃排空(gastric emptying)顯著延遲,食物滯留導致早飽與嘔吐。理解這一點,治療邏輯就清楚了:促進排空的藥物(如作用於運動的促動劑)、少量多餐、降低餐點脂肪與纖維(兩者都會延緩胃排空),都是針對「運動」而非「酸」或「酵素」的策略。這個案例凸顯進階觀念——許多消化問題的根源不在化學消化,而在運動與神經控制。(本文為教育用途,不構成個人醫療建議。)
第二大腦的內部結構:腸神經系統如何「不請示大腦」就運作
入門篇提到腸神經系統(enteric nervous system, ENS)被稱為「第二大腦」、含有數億個神經元。進階篇要看清楚它為什麼能獨立運作,以及它與中樞神經的真實關係。
ENS 由兩個神經叢構成:位於兩層肌肉之間、主管運動的肌間神經叢(myenteric plexus, Auerbach's plexus),以及位於黏膜下層、主管分泌與局部血流的黏膜下神經叢(submucosal plexus, Meissner's plexus)。它擁有完整的反射弧——感覺神經元(偵測腸腔的張力與化學成分)、中間神經元(整合運算)、運動神經元(驅動平滑肌與腺體)——這正是它能在離體(把一段腸子取出培養)狀態下,仍自行產生蠕動反射的原因。
最經典的內建程式是蠕動反射(peristaltic reflex):腸腔內某處被食團撐開時,ENS 會同時命令口側收縮、肛側舒張,形成一個把內容物往下游推的行進波。口側的收縮靠興奮性神經元釋放乙醯膽鹼與 P 物質,肛側的舒張靠抑制性神經元釋放一氧化氮與血管活性腸肽(VIP)。這種「前舒後縮」的協調,是 ENS 自己算出來的,中樞只在背景做調節。
理解 ENS 的自主性,也就理解了一個重大的臨床原型——先天性巨結腸症(Hirschsprung disease)。這類嬰兒因為胚胎期神經嵴細胞遷移失敗,遠端結腸缺乏神經節細胞(aganglionosis),那一段腸子失去抑制性神經元、無法舒張,於是持續痙攣收縮、糞便堆積在上游,造成嚴重便祕與腸阻塞。它揭示了一個容易被忽略的原則:腸道的舒張和收縮一樣,都是被「主動驅動」的;少了負責「放鬆」的神經元,腸子並不會默認打開,反而會卡死。
腸—腦軸的雙向高速公路:不只是「壓力會拉肚子」
入門篇與多數科普把腸—腦軸(gut-brain axis)講成「情緒影響腸胃」。進階視角要強調它是雙向、且大半是由下往上的。
從解剖看,連接腸與腦的主幹是迷走神經(vagus nerve),但有個常被忽略的數字:迷走神經中約 80–90% 是傳入(afferent)纖維,也就是把腸道訊息往上送到腦,而非腦往下指揮腸。這意味著腸道更像是一個不斷向中樞「回報現場狀況」的感測陣列。腸道內分泌細胞(enteroendocrine cells)能感知管腔內的營養與菌群代謝物,釋放訊號分子;近年研究甚至發現一類神經足細胞(neuropod cells)能與迷走神經形成類突觸連結,在毫秒尺度上把腸腔的營養訊息直接傳給腦——這比過去認為的「荷爾蒙慢速擴散」快得多。
這條軸線還有第三條路徑:腸道菌群的代謝產物。菌群發酵纖維產生的短鏈脂肪酸、由色胺酸(tryptophan)衍生的代謝物、以及菌群影響的神經傳導物質前驅物,都能調節免疫、迷走訊號甚至中樞功能。一個近年備受關注的線索是帕金森氏症:部分證據顯示,異常摺疊的 α-突觸核蛋白(α-synuclein)可能先出現在腸神經系統,再沿迷走神經「向上」傳播到腦幹——亦即某些神經退化疾病的起點,或許在腸而不在腦。這個假說仍在驗證中,但它徹底翻轉了「腦指揮腸」的單向直覺。
膽酸不只是清潔劑:腸—肝循環與作為訊號分子的膽汁
入門篇把膽汁(bile)的角色定位為「乳化脂肪的界面活性劑」。這只說對了一半。進階篇要補上兩件大事:腸—肝循環的回收效率,以及膽酸作為荷爾蒙的訊號功能。
先談回收。膽鹽(bile salts)並非用過即棄。它們在迴腸末端被頂端鈉依賴性膽酸轉運蛋白(ASBT)主動回收,經肝門靜脈送回肝臟再利用,這個循環稱為腸—肝循環(enterohepatic circulation)。其效率高得驚人:人體膽酸池(bile acid pool)只有約 2–4 克,卻能在一餐之間循環數次、一天總共周轉約 6 至 8 次,相當於每天「搬運」20–30 克膽酸卻幾乎不耗損。肝臟每天只需新合成約 0.5 克來補足隨糞便流失的少量。這也解釋了一個臨床現象:若迴腸被切除或發炎(如克隆氏症 Crohn's disease),膽酸回收失敗、大量膽酸進入大腸刺激分泌,會造成膽酸性腹瀉(bile acid diarrhea);同時膽酸池縮小、膽汁中膽固醇相對過飽和,膽結石風險上升。
更前沿的觀念是:膽酸是訊號分子,不只是清潔劑。膽酸能活化核受體FXR(farnesoid X receptor)與細胞膜受體TGR5。透過 FXR,腸道偵測到膽酸後會分泌纖維母細胞生長因子 19(FGF19),回頭告訴肝臟「膽酸夠了,別再合成」,形成一個跨器官的負回饋;透過 TGR5,膽酸還能促進腸道 L 細胞釋放 GLP-1、調節能量消耗與葡萄糖代謝。這就是為什麼膽酸—FXR—FGF19 軸成為治療代謝相關脂肪肝(MASLD)與膽汁淤積的藥物標的。把膽汁僅僅看成「洗油的肥皂水」,會錯過它在血糖、脂質與肝臟健康中的調控角色。
動手試試:用「回收率」估算為什麼少量膽酸夠用
給自己一個量化練習。已知膽酸池約 3 克、每天周轉約 8 次,估算一天經由腸道「處理」的膽酸總量,並與每日新合成量比較。
簡單推算:每天通過量 ≈ 3 克 × 8 次 = 24 克。而肝臟每天只新合成約 0.5 克(補足糞便流失)。兩者相除,回收率 ≈ (24 − 0.5) / 24 ≈ 98%。這個數字告訴你:身體不是靠「大量製造」來供應膽汁,而是靠「近乎無損的回收」。一旦回收環節(迴腸 ASBT)出問題,即使肝臟拚命合成也補不回 24 克的周轉量——這正是迴腸疾病導致脂肪吸收不良與腹瀉的數量級原因。用一個簡單的乘除,你就把解剖位置(迴腸末端)、轉運蛋白(ASBT)與臨床後果(膽酸性腹瀉)串成了一條因果鏈。
腸黏膜屏障:一道「選擇性開放」的免疫前線
入門篇提過腸道黏膜要「兼具吸收與屏障」。進階篇要拆解這道屏障究竟由什麼構成,以及它如何在「讓營養進來」與「擋細菌出去」之間動態取捨。
腸屏障至少有三層。物理層是單層上皮細胞與細胞之間的緊密連接(tight junctions),其通透性由 claudin、occludin 等蛋白動態調節——它不是一道死牆,而是可被訊號(如 zonulin、發炎細胞激素)開關的「閘門」。化學層包括杯狀細胞(goblet cells)分泌的黏液層,以及潘氏細胞(Paneth cells)分泌的抗菌肽(defensins),在上皮表面築起一道讓細菌難以貼近的緩衝帶。免疫層則由腸道相關淋巴組織(GALT)、分泌型 IgA 與大量常駐免疫細胞組成,持續監測管腔。
這道屏障最迷人的地方在於它的選擇性矛盾:它必須對食物抗原與共生菌「容忍(tolerance)」,又要對病原體「警戒」。負責拿捏分寸的是派氏結(Peyer's patches)上的 M 細胞與其下的免疫細胞,以及一群維持免疫耐受的調節性 T 細胞(Treg)——而 Treg 的誘導,部分正是靠前面提到的菌群代謝物丁酸(butyrate)。當這套平衡被打破、屏障通透性異常升高(俗稱「腸漏」的學術版本是 increased intestinal permeability),管腔內容物滲入引發慢性免疫活化,被認為與發炎性腸道疾病、乳糜瀉(coeliac disease)等多種狀況有關。這層理解把消化道從「營養吸收管」升級為「全身最大的免疫介面」——人體約七成的免疫細胞駐紮於此並非偶然。
重點回顧
- 消化道是有自主節律的器官:空腹時靠移行性運動複合波(MMC)清掃腸道,並非進食才工作;ICC 起搏細胞產生的慢波決定各段腸道的收縮頻率與方向性。
- 腸神經系統(ENS)擁有完整反射弧,能獨立執行「前舒後縮」的蠕動反射;舒張與收縮一樣都需要被主動驅動,缺乏抑制性神經元(如 Hirschsprung disease)反而造成腸子痙攣卡死。
- 腸—腦軸是雙向且以「由下往上」為主:迷走神經約八至九成是傳入纖維,腸道更像不斷向腦回報的感測陣列,菌群代謝物與 α-synuclein 假說延伸出神經退化疾病可能起於腸的觀點。
- 膽汁靠腸—肝循環近乎無損回收(回收率約 98%);膽酸更是活化 FXR/TGR5 的訊號分子,透過 FGF19 與 GLP-1 參與血糖、脂質與肝臟代謝調控。
- 腸黏膜屏障由物理(緊密連接)、化學(黏液與抗菌肽)、免疫(GALT、IgA、Treg)三層構成,需在「容忍食物與共生菌」與「警戒病原」間動態取捨,是全身最大的免疫前線。
深入探討(研究所視角)
把消化系統重新理解為「運動—神經—內分泌—免疫」的整合器官後,幾條當代研究主線會自然浮現。
運動電生理與 ICC 的可塑性:糖尿病性胃輕癱中的 ICC 流失是可逆還是不可逆?近年動物研究顯示 ICC 數量受幹細胞因子(SCF)與胰島素/IGF-1 訊號維持,這為「修復起搏網路」而非僅「症狀治療」打開了新方向。閱讀這類文獻時,要留意胃排空閃爍掃描(gastric emptying scintigraphy)等測量工具的標準化問題——終點指標若不一致,跨研究比較會失真。
膽酸訊號與代謝藥理學:FXR 促效劑(如 obeticholic acid)與 FGF19 類似物正被開發用於膽汁淤積與代謝性肝病,但 FXR 在不同組織(肝、腸)作用方向不同,全身性活化的副作用(如搔癢、血脂變化)是臨床轉譯的核心挑戰。這提醒一個方法學要點:當一個受體在多器官表達,「組織選擇性藥物」與「給藥途徑」往往比「受體本身」更決定療效與安全性。
腸—腦軸的因果鑑定:菌群與神經精神疾病的關聯多來自觀察性或動物研究,從「相關」走到「因果」極為困難。理想設計需要無菌(germ-free)動物的菌群移植、人體的隨機介入(如特定益生菌或糞菌移植 RCT),並輔以代謝體學鎖定真正起作用的分子。閱讀此領域時,對「小鼠結果直接外推到人」與「橫斷面菌群差異等於致病」兩種跳躍,要保持高度警覺。
屏障免疫與精準介入:把「腸漏」從通俗概念變成可測量、可介入的科學,需要可靠的通透性生物標記與機制證據。短鏈脂肪酸誘導 Treg、膳食纖維塑形菌群與屏障的研究,正試圖把營養介入建立在分子機制之上。嚴謹的判讀應同時要求機制證據(細胞與動物模型)與臨床終點(症狀、發炎指標、復發率)的一致——兩者並置,才是把消化生理推向治療的可信路徑。